Paper 11
The Effect of 6-Benzyl Amino Purine on Indole Acetic Acid Mediated Molecular Events Leading to the New Root
Formation.
G. R. KANTHARAJ* and G. PADMANABHAN**
* Dept. of Botany, The National Collage, Bangalore – 560 004, India. (Correspondence address)
Dept. of Biochemistry, Indian Institute of Science, Bangalore - 560 012,
India
INTRODUCTION
Plant hormones are capable of multifarious functions. At the morphogenetic level, they are capable of inducing both growth and differentiation. It is rather difficult to discern and segregate the biochemical effects of the hormones meant for growth and differentiation. Thus, IBA enhances protein and RNA synthesis in the hypocotyls tissue of P. vulgaris, which perhaps are necessary for both cell division and differentiation. General inhibitors of protein synthesis such as cycloheximide may not be very useful in identifying the synthesis of specific macromolecules which are probably essential for the differentiation process. It was felt that studies on hormone-hormone interaction may help in identifying the specific events involved in differentiation. Cytokinin is known to inhibit auxin-mediated changes in growth and RNA metabolism in soybean hypocotyls [100, 101] and therefore its effects on IBA-induced root initiation were studied.
MATERIALS AND METHODS
Materials
6-Benzylaminopurine (BAP) was purchased from Sigma Chemicals Company, U.S.A. All other chemicals used were of analytical grade.
Methods:
Effect of 6-Benzylaminopurine on IBA-mediated new root formation:
Excised hypocotyls segments (8-10 cm length) were incubated in Hoagland’s nutrient medium containing 30 mg/ml of IBA and increasing concentrations of BAP (10 ng – 1 mg/ml), for 30 minute. After incubation, segments were washed in running water and transferred to fresh medium. At the end of 72 hr, root production was observed.
Anatomical studies:
Hormone treated hypocotyls segments were cut into 1 cm segments, fixed, embedded in paraffin was and processed for micro slide preparations as described in experimental methods of Chapter I.
RNA and protein synthesis:
Hypocotyl segments, after hormone treatment were exposed to radioactive precursors [32 P] – orthophosphate for RNA and [14 C]-chlorella protein hydrolysate for protein) for 30 minutes. Then the tissue was processed for the rate of protein synthesis and RNA synthesis as described in experimental methods is Chapter I.
Isolation of total RNA and translation of the same in a cell-free system derived from wheat germ:
The procedures used for isolating total RNA and in vitro translation are the same as those described in Chapters I and II.
Analysis of membrane proteins in SDS-gels:
After treating the hypocotyls tissue with phytohormones, membrane proteins were isolated and electrophoresed on SDS-gels as described in Chapter I and II.
Flurographic analysis of in vivo labeled proteins:
The excised hypocotyls tissue was treated with IBA, BAP or IBA + BAP (IBA, 30 mg/ml; BAP, 10 mg/ml) for 30 minutes. Then the segments were transferred to a fresh medium. After 34 hours of incubation, hypocotyls sections were exposed to 25 mCi of [14 C]-chlorella protein hydrolysate for 2 hr. After labeling, the hypocotyls segments were homogenized in 0.02M sodium phosphate buffer (pH 7.0) containing 0.25M sucrose, and 10 mM PMSF in a pre-cooled all-glass pestle and mortar. The homogenate was centrifuged at 1500 rpm for 10 minutes and the supernatant was carefully removed. The supernatant protein (250 mg) was dissociated in presence of 2% SDS and electrophoresed on 8% SDS-gels and fluorographed as described in Chapter III.
Analysis of in vitro translational protein products on SDS-PAGE:
Total RNA isolated from control and hormone-treated hypocotyls was translated in wheat germ cell-free system in presence of [3 H] – leucine. The labeled protein product was analysed on SDS-gels as described in Chapter II. After electrophoresis, the gels were sliced, digested with H202 and counted for radioactivity.
RESULTS
Effect of 6-Benzylaminopurine on IBA-induced new root formation:
To begin with the effect of 6-benzylaminopurine on new root formation was studied in the presence and absence of IBA. The results (Fig.27) indicate that the cytokinin by itself does not induce root formation in the hypocotyls tissue. At low concentrations (10 ng /, l) it has a synergistic effect on the root formation induced by IBA. At higher concentrations, BAP inhibits the IBA mediated effects and at concentration of 1 mg/ml and above, the cytokinin completely inhibits root initiation.
Determination of BAP-sensitive phase during IBA-induced adventitious root formation:
The inhibitory effects of cytokinin were studied as a function of the time of addition of the hormone. The results
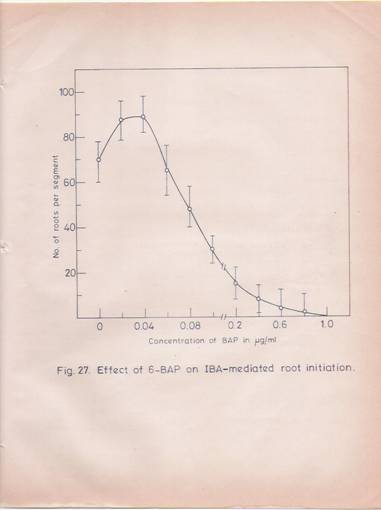
Fig. 27: Effect of 6-Benzylamino purine on Indolebutyric acid-mediated root initiation.
Hypocotyl segments (8-10 cm length) which were pretreated with IBA (30 mg/ml for 30 min) were placed in Hoagland’s nutrient medium in flasks containing increased amounts of BAP. After incubating with cytokinin for 30 min, the segments were washed thoroughly and transferred to fresh basal medium. At the end of 72 hr the roots were counted. The number of roots produced is presented in the Figure as the average of 10 segments with standard deviations.
Presented in Fig. 28. suggests that, when hypocotyls are incubated with cytokinin after 36 hr of the IBA treatment the BAP fails to inhibit root formation. But, cytokinin, when added at time points prior to 36 hr, abolishes IBA mediated root formation.
Anatomical studies:
The anatomical changes taking place in the hypocotyl tissue after treatment with the cytokinin at inhibitory concentrations are shown in Fig. 29. BAF alone does not affect even general cell division, but there is no organization of cells into root primordial. In BAP + IBA treated hypocotyls an increased cell-division all-round the vascular bundle is observed, but root primordial is totally absent.
.
Effect of BAP on IBA-induced protein and RNA synthesis:
To localize the effects of BAP at the biochemical level, the effects of the hormone on protein and RNA synthesis in the hypocotyls segments were studied both with and without IBA treatments. Cytokinin alone enhances protein synthesis as early as 30 min after the hormone treatment. This effect is similar to that observed with IBA. However, cytokinin + IBA treatment does not produce an additive effect on the rate of protein synthesis (Table 15). The enhanced rate of protein synthesis is maintained in the hypocotyls treated with IBA, BAP and IBA + BAP for a period of 24 hr. The effect of

Fig. 28: Effect of Benzyl amino purine on IBA induced root initiation at various stages of development.
Hypocotyl segments (8-10 on length) were treated with IBA (30 mg/ml) for 30 min and then transferred to the basal medium. At various time periods, BAP (10 mg/ml) was added. After incubating for 30 min, segments were washed with water and incubated in fresh nutrient solution. After every 24 hr the medium was charged. At the end of 72 hr roots produced were counted. The number of roots per segment was given as the average of 10 segments with standard deviations.

Fig. 29: Microphotographs of transverse sections of hypocotyls segments treated with IBA, BAP and IBA + BAP.
(A) Transverse section of hypocotyls at 0 hr. X 200.
(B) Transverse section of IBA-treated hypocotyls at 72 hr showing well advanced root primordial through cortex. X 200.
(C) Transverse section of BAP-treated hypocotyls at 72 hr showing few cell divisions around the vasculature. Root primordials are not found. X 200.
(D) Transverse section of IBA + BAP-treated hypocotyls showing numerous cell divisions all round vascular bundles. Root primordial is conspicuously absent. X 200.
Table 15: Effect of BAP on IBA mediated protein synthesis in the hypocotyls segments.
|
Treatment |
Time |
A Amino acid Pool (cpm /mg) |
B Protein
(cpm /mg) |
B/A Protein / amino acid Pool |
|
Control IBA BAP IBA + BAP |
30 min |
72.0 75.3 81.8 69.7 |
5.5 12.0 10.7 7.6 |
0.07 0.15 0.13 0.11 |
|
Control IBA BAP IBA + BAP |
3 hr |
69.1 63.8 71.1 67.6 |
5.7 12.4 8.5 8.5 |
0.08 0.19 0.11 0.12 |
|
Control IBA BAP IBA + BAP |
6 hr |
65.8 59.0 78.1 74.4 |
4.3 11.8 10.2 10.8 |
0.06 0.20 0.13 0.14 |
|
Control IBA BAP IBA + BAP |
12 hr |
71.4 41.2 41.9 49.0 |
6.3 12.0 10.8 14.7 |
0.08 0.29 0.25 0.30 |
|
Control IBA BAP IBA + BAP |
24 hr |
75.9 60.4 61.1 61.6 |
8.5 28.2 24.8 29.5 |
0.11 0.46 0.40 0.47 |
Table 15 (contd.)
Hypocotyl tissue (4 cm length, 1 gm) was treated with IBA (30 mg/ml), BAF (30 mg/ml) or IBA + BAF (30 mg/ml each) for 30 minutes. After washing with water, the segments were transferred to fresh medium and at various time points, the tissue was incubated with 2.5 mCi of [14 C]-chlorella protein hydrolysate for 30 minutes. Then the hypocotyls were washed and processed for estimating free amino acid pool and proteins as described in Materials and Methods in Chapter I.
Cytokinin treatment on total, as well as poly (A)-containing RNA synthesis is significant (Table 16). BAP treatment enhances the rate of RNA synthesis after a lag period of about 12 hr, whereas IBA treatment results in enhanced RNA synthesis as early as 2 hr after the hormone treatment. In IBA + BAP treatment, the pattern of RNA synthesis follows that of BAP treatment rather than that of IBA treatment.
Effect of 6-Benzylaminopurine on protein distribution among soluble and membrane fractions:
Since, IBA treatment was found to result in enhanced membrane protein content (Chapter III), the effect of cytokinin was examined in this regard. At 24, 48 and 72 hr after the hormone treatment, both IBA and cytokinin enhance the membrane protein content of the hypocotyls tissue. However, the effects are not additive (Table 17).
SDS-Gel Analysis of membrane proteins obtained from IBA, BAP and IBA + BAP-treated hypocotyls:
Subsequently, the effect of cytokinin on tubulin formation was studied. The SDS-gel analysis of the membrane proteins (Fig. 30) indicate, that tubulin is formed under conditions of IBA, CYTOKININ AS WELL AS IBA + BAP treatments.
Table 16: Effect of BAP on IBA – induced RNA synthesis in hypocotyls tissue.
|
Treatment |
Time |
Nucleotide pool (cpm/A260) A |
Total RNA
(cpm/A260) B |
B/A |
Poly (A) ± RNA (cpm/30 A260 units of total RNA |
|
Control |
30 min |
12200 |
1650 |
0.13 |
340 |
|
|
2 hr |
12670 |
1550 |
0.12 |
360 |
|
|
4 hr |
13070 |
1720 |
0.13 |
350 |
|
|
12 hr |
14650 |
2750 |
0.18 |
425 |
|
|
24 hr |
14360 |
2600 |
0.18 |
485 |
|
|
|
|
|
|
|
|
IBA |
30 min |
12000 |
1450 |
0.12 |
310 |
|
|
2 hr |
12480 |
2770 |
0.20 |
510 |
|
|
4 hr |
12140 |
2940 |
0.24 |
700 |
|
|
12 hr |
14480 |
4250 |
0.29 |
1150 |
|
|
24 hr |
14100 |
4800 |
0.33 |
1380 |
|
|
|
|
|
|
|
|
BAP |
30 min |
13370 |
1530 |
0.11 |
330 |
|
|
2 hr |
13200 |
1650 |
0.12 |
390 |
|
|
4 hr |
13790 |
2070 |
0.15 |
367 |
|
|
12 hr |
14870 |
4050 |
0.27 |
858 |
|
|
24 hr |
14380 |
4100 |
0.28 |
925 |
|
|
|
|
|
|
|
|
IBA+BAP |
30 min |
11640 |
1450 |
0.12 |
340 |
|
|
2 hr |
12660 |
1650 |
0.13 |
375 |
|
|
4 hr |
11660 |
2250 |
0.19 |
477 |
|
|
12 hr |
14280 |
4300 |
0.30 |
1080 |
|
|
24 hr |
13960 |
5800 |
0.41 |
2100 |
|
|
|
|
|
|
|
|
|
|
|
|
|
.. contd. |
Table 16 (contd.)
Hypocotyl segments (4 cm length, 20 gm) were treated with IBA, BAP or IBA + BAP (30 mg/ml each) for 30 minutes. After washing with water, the segments were transferred to fresh basal medium and at different time periods of development the segments were incubated in 5 ml of nutrient medium containing 100 mCi of [32 P] –orthophosphate (carrier free) for 30 minutes. Then the segments were washed and processed for measuring total RNA and poly (A)-containing RNA synthesis as described in experimental part in Chapter I.
Table 17: Effect of IBA, BAP and IBA + BAP on soluble and membrane protein contents.
|
Treatment |
Time |
Soluble protein ( mg ) |
Membrane protein ( mg ) |
Membrane / soluble protein |
|
Control |
0 hr |
27.6 |
2.30 |
0.083±0.028 |
|
|
24 hr |
21.0 |
1.66 |
0.079±0.023 |
|
|
48 hr |
22.3 |
1.70 |
0.076±0.020 |
|
|
72 hr |
22.0 |
1.60 |
0.072±0.021 |
|
|
|
|
|
|
|
IBA |
0 hr |
27.6 |
2.30 |
0.083±0.028 |
|
|
24 hr |
18.6 |
1.88 |
0.100±0.030 |
|
|
48 hr |
23.3 |
2.80 |
0.120±0.028 |
|
|
72 hr |
17.6 |
2.50 |
0.140±0.016 |
|
|
|
|
|
|
|
BAP |
0 hr |
27.6 |
2.30 |
0.083±0.028 |
|
|
24 hr |
22.0 |
2.05 |
0.090±0.032 |
|
|
48 hr |
21.0 |
2.16 |
0.100±0.022 |
|
|
72 hr |
21.0 |
2.36 |
0.110±0.031 |
|
|
|
|
|
|
|
IBA+BAP |
0 hr |
27.6 |
2.30 |
0.083±0.028 |
|
|
24 hr |
36.6 |
3.40 |
0.090±.021 |
|
|
48 hr |
38.2 |
4.00 |
0.100±0.030 |
|
|
72 hr |
39.1 |
4.90 |
0.120±0.028 |
|
|
|
|
|
|
Soluble and membrane fractions were prepared from the treated and control hypocotyls as explained in Materials and methods. Protein was estimated by the method of Lowry et al. [221] and the data are presented as a ratio between membrane and soluble proteins. The values represent the mean ±S.D. obtained from three independent experiments.

Fig. 30: SDS-slab gel electrophoresis of membrane proteins.
The experimental procedure is described in Materials and Methods. Membrane protein (250 mg) was electrophoresed.
Lanes 1, 6 and 11 are Standard Proteins.
Lanes 2,3,4 & 5; 7,8,9 & 10 and 12, 13, 14 & 15 depict protein from membrane fractions of control, IBA, BAP and IBA + BAP at 24 hr and 72 hr respectively.
Lane 16 shows purified rat brain tubulin.
Protein synthesis in vivo, in response to IBA, BAP and IBA + BAP treatment:
It was of interest to examine the total protein profiles
Of the hormone treated hypocotyls tissues and in order to look for changes in minor proteins, the segments were labeled with [14C]-chlorella protein hydrolysate and the fluorograph of the labeled proteins after 36 hr of hormone treatment are shown in Fig. 31. Qualitative changes in the total protein profiles are not apparent. However, in IBA + BAP treated hypocotyls the labeling of certain high molecular weight proteins is significantly decreased.
Quantification of mRNA by in vitro translation in a cell-free system derived from wheat germ and analysis of translated products by SDS-PAGE:
An attempt was also made to examine whether the effects of BAP on the pattern of protein synthesis is also reflected at the level of the messenger RNA populations. For this purpose, total RNA was isolated from control, IBA, BAP and IBA + BAP treated hypocotyls and the RNA was translated in wheat germ cell-free system. The result presented in table is messenger RNA activity at 24 hr and 48 hr after both hormone treatments. An analysis of the labeled protein products is presented in Fig 32. It can be seen that the conspicuous effect of BAP treatment is a decrease in the messenger populations coding for high molecular weight proteins and increases in the messenger content for low molecular weight proteins. This effect is more pronounced in the case of RNA preparations isolated from BAP and BAP + IBA treated hypocotyls.

Fig. 31: Flurographic analysis of total proteins synthesized in the hypocotyls segments treated with IBA, BAP and IBA + BAP for 36 ht.
Hypocotyl segments (2 g) which were already treated with IBA, BAP OR A +BAP for 34 hr were transferred into 2 ml nutrient medium containing 40 mCi of [14C]-chlorella protein hydrolysate. After incubating for 2 hours, the segments were washed with water and total proteins were obtained as described in Chapter I After electrophoresis the gels were stained, impregnated with PPO, dried and fluorographed for 15 days as described in chapter III.
Lane 1 – control.
Lane 2, 3 and 4 – proteins from IBA, BAP and IBA + BAP treated segments.
Table 18: Quantification of mRNA by in vitro translation in cell free-system obtained from wheat germ.
Treatment Time [3H]-Leucine incorporated into TCA
Precipitable protein (cpm/A260 unit of RNA
Translated)
__________________________________
Experiment I Experiment II Experiment III
Control 12 hr 46640 49500 48000
24 hr 45740 54300 52300
48 hr 51660 46000 48000
IBA 12 hr 75780 80120 78650
24 hr 136170 142000 138200
48 hr 165300 176000 169300
BPA 12 hr 72500 70150 70100
24 hr 160580 150000 155300
48 hr 154660 178000 170000
IBA+BAP 12 hr 88670 89720 87680
24 hr 164250 168000 167000
48 hr 228000 268000 258660
The total RNA was extracted from control, IBA, BAP and IBA + BAP-treated segments and total RNA from each sample was translated in cell-free extracts obtained from wheat germ, as described in chapter II. Results of two independent experiments are given. Each value is the mean of two replicates.

Fig. 32: Radioactivity profile of total RNA translated products in the cell-free system derived from wheat germ.
Equal amounts of total RNA from each sample were translated and the entire protein product was electrophoresed on SDS-gel. The gels were sliced (1 mm) and counted.
RNA translated protein products obtained from:
(A) Control
(B) IBA treated (_ _ _ _ _ _) and IBA + BAP treated (_________) for 24 hr.
(C) IBA treated (_ _ _ _ _ _) and IBA + BAP treated (_________) for 48 hr.
(D) IBA treated (_ _ _ _ _ _) and BAP treated (________) for 24 hr.
(E) IBA treated (_ _ _ _ _ _) and BAP treated (________) for 48 hr.
(F)
DISCUSSION
In the present study, an attempt has been made to specifically identify the biochemical events leading to adventitious root initiation brought about by IBA treatment. Since the hormone causes both cell elongation and differentiation the gross changes in RNA and protein synthesis would account for both the processes. The results presented in chapter II have indicated that cycloheximide treatment abolishes the IBA-induced very early protein synthesis and delays RNA synthesis. There is a striking inhibition of IBA-induced root formation. Cycloheximide being a general inhibitor of protein synthesis, a more specific antagonist for IBA action was looked for.
Synergistic as well as antagonistic interactions (Fig .1) between plant hormones are well known [100,101]. In the present study, the cytokinin (BAP) has been found to antagonize the root inducing effects of IBA at concentrations greater than 1 mg/ml. At lower concentrations, it has synergistic effects in terms of root production. BAP treatment by itself does not induce adventitious root initiation. AT concentrations inhibiting IBA-Induced new root formation, BAP enhances protein and RNA synthesis. These effects are similar to those obtained with IBA except that the increase in RNA synthesis due to BAP treatment takes place after a lag of about 12 hr. The effects obtained with IBA + BAP treatment are not additive and the pattern is similar to that obtained with BAP treatment. It appears that some of the early messengers and early proteins synthesized in response to IBA are inhibited significantly in the case of BAP treatment. Such a situation is indicated also in the case of IBA+BAP treatment, where the hypocotyls tissue proteins labeled in vitro as well as the labeled proteins synthesized in vitro in a cell-free system show decreased levels of certain high molecular weight proteins and perhaps this may prevent the cells being committed to new root formation. However, in IBA+BAP treatment, cell division all-round vascular bundle is greatly enhanced but without the organization of any root primordial. On the contrary, in the case of BAP treatment alone, even the increase in cell number is inhibited, thereby indicating that cytokinin requires auxin in effecting cell divisions.
The formation of tubulin, an important component of the microtubule and cytoskeletal assemblage, which play a vital role in determining cell shape, morphogenesis and differentiation, is not affected by BAP treatment. Thus, while tubulin formation in response to IBA treatment is a vital event in adventitious root initiation, the results obtained with BAP indicate that there are other factors which regulate tubulin assembly into specific cytoskeletal structures. It appears possible that BAP may be interfering with the formation of specific proteins, which regulate the formation of cytoskeletal assemblage leading to cell polarity fixation and the formation of root primordial. In this context it may be pointed out that accessory proteins such as MAPs have been implicated to play a role in determining the structures to be formed in vivo [240,243]. It is possible that the high molecular weight proteins whose synthesis is inhibited by BAP treatment along with tubulin may be the proteins responsible for the differentiation process. However, it is clear that BAP, in the presence of IBA permits the synthesis of tubulin as well as proteins necessary for cell division to proceed.
SUMMARY
1. 6-Benzylamino purine exhibits synergistic effect on IBA-induced new root formation at the concentration of 10 ng/ml but at higher concentration of 1 mg/ml and above, it inhibits IBA mediated root initiation.
2. Cytokinin-sensitive phase for IBA induced root formation is 36 hr.
3. BAP in the presence of IBA enhances cell division all-round the vascular bundle in the hypocotyls segment, but without any root primordial organization. BAP by it self hardly has any effect either on root initiation or on cell division.
4. Protein synthesis is enhanced as early as 30 minutes after BAP treatment. Similar effect is also found in cytokinin + IBA treated segments, but there is no additive effect on protein synthesis.
5. While BAP treatment enhances the rate of total as well as poly(A)-containing RNA synthesis after a lag period of 12 hr, IBA increases RNA synthesis within 2 hr of treatment, but RNA synthesis In BPA+IBA-treated segments follows the same pattern as that of BAP treated segments.
6. IBA, BAP and BAP+ IBA treatments cause an increase in the content of membrane proteins. They also induce tubulin synthesis.
7. Radioactivity profiles of proteins labeled in vivo show a significant decrease in high molecular weight proteins in BAP and BAP + IBA treated segments.
8. Analysis of translation products of total RNA in cell free system derived from wheat germ shows that BAP treatment causes a decrease in messenger population coding for high molecular weight proteins. This effect is also seen in IBA + BAP treated segments.