Paper 5
TUBULIN SYNTHESIS AND AUXIN-INDUCED ROOT INITIATION IN PHASLOLUS
G. R. KANTHRAJ, * S. MAHADEVAN and G. PADMANABHAN
*Department of Botany, National College, Bangalore 560 00 Indian Institute of Science, Bangalore, India
(Revised received 15 June 1984)
![]()
Abstract: The auxin-induced formation of roots in the hypocotyls of Phaseolus vulgaris can be prevented by treatment with Actinomycin D, colchicine or cytochalasin B if applied within 40 hr of initiation. Shortly after auxin pretreatment, there is an increase in translatable messenger RNA activity. Analysis of the labeled cell-free products indicates, among other changes, a striking increase in a protein co-migrating with tubulin, in the case of RNA isolated from Indole Butyric Acid (IBA) pretreated hypocotyls. An increase in tubulin content in vivo can also be demonstrated on the basis of SDS polyacrylamide gel analysis of membrane proteins and Functional assays for tubulin polymerization. An increase in the synthesis of tubulin in vivo can also be demonstrated after IBA pretreatment. In addition, the auxin is also able to promote tubulin polymerization when added in vivo. It is suggested among others that tubulin synthesis and microtubule assembly are some of the early events in auxin-mediated root differentiation.
INTRODUCTION
Among the factors, which regulate adventitious root formation, auxin is of primary importance [1]. Excised or intact hypocotyls and epicotyls segments undergo redifferentiation in response to auxin treatment, leading to the formation of root primordial. The morphological and biochemical events underlying this process have been analyzed. In auxin-treated pea epicotyls or soybean hypocotyls, the parenchymatous cells in the cortex appear swollen and exhibit few cell divisions 24 hr later. After 48 hr, the number of cell divisions increases and disintegration of many cortical cells sets in, leaving lacunae. Root primordials originate from cambial cells near vascular tissues and become recognizable after 3 days. The primordial cells continue to divide and occupy the lacunae. In 4-5 days, the new roots break through the epidermal layers of the auxin-treated segments [2-4].
Several biochemical changes have been recorded during the process of auxin-induced new root formation. These include an increase in RNA, DNA and protein contents, a shift in polysomes, and an increase in the labeling of rRNA, tRNA and DNA-like RNA and ribosomes [4-7]. After 3 days of auxin treatment, a significant increase in the activities of cellulase, pectin esterase and b (1 - 3) gluconase have been reported [3]. The increase in cellulase activity is totally abolished if the segments are treated with Actinomycin D or Azaguanine or Puromycin together with auxin [4].
The present study is an attempt to identify specific auxin-induced, early macromolecular changes in the hypocotyls of Phaseolus vulgaris during new root formation induced by auxin treatment.
RESULTS
In our laboratory, it was shown that auxin pretreatment of hypocotyls obtained from Phaseolus vulgaris seedlings leads to an increase in Actinomycin D insensitive protein synthesis within 30 min. Pretreatment of hypocotyls segments with cycloheximide together with auxin greatly delays and diminishes the formation of new roots. In ca 2 hr after auxin pretreatment, there is an increase in the synthesis of RNA and Actinomycin D sensitive protein synthesis [8].
In the present study, the effects of Actinomycin D, Cytochalasin B and Colchicine on root production by auxin-pretreated hypocotyls were tested. All three drugs completely inhibited the emergence of root primordials when added at any time during the first 36-40 hr after IBA pretreatment, but permitted almost normal root formation when added after this time period (data not presented). In view of the earlier observations on the IBA mediated increase in total RNA synthesis and RNA dependent protein synthesis (I), and the fact that Actinomycin D and the other two drugs would interfere with transcription and microtubule integrity respectively. The effects of auxin pretreatment on messenger RNA activity and the nature of the protein products synthesized were studied.
The total and poly (A)-containing RNAs were isolated from untreated and IBA-pretreated hypocotyls segments at different periods of time. The messenger activity of total RNA was tested in the wheat germ cell-free system. The messenger RNA content as assayed in terms of translatable activity increases by 2- to 4-fold over a period of 6-48 hr after IBA pretreatment (Fig. I). It is also seen that at equivalent concentrations, poly (A)-containing RNA preparations from IBA-pretreated hypocotyls at different periods of time uniformly exhibit 1.5-fold higher transnational efficiency than the RNA preparations from control segments at the corresponding time periods (Table 1). It thus appears that auxin pretreatment enhances messenger RNA content as well as its transnational efficiency.
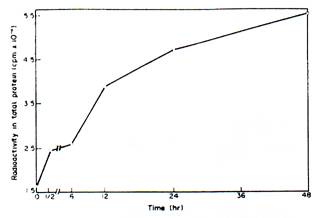
Fig. 1. Effect of IBA pretreatment on the translatable activity of total RNA from Phaseolus vulgarism. Total RNA was isolated from the hypocotyls at various adage of development after IHA pretreatment, and 30 pg of RNA from each sample was translated using the wheat germ all-free system in a total volume of 100 ml.
Other experimental details art given in the text.
![]() Table
1. Effect of IBA pretreatment on the translatable activity of poly (A)-
Table
1. Effect of IBA pretreatment on the translatable activity of poly (A)-
containing RNA from Phaseolus vulgaris
[3H]Leucine incorporated into TCA – precipitable proteins (cpm / 5m g of poly (A) -RNA
![]() RNA Time
RNA Time
Source IBA
Source Pretreatment I II III
Control* All the time 47200 39400 46000
Intervals
IBA 30 min 62000 62500 60500
IBA 6 hr 64800 64300 63500
IBA 12 hr - 70000 68000
![]()
Poly (A) containing RNA was isolated from the hypocotyls at various stages of development after IBA pretreatment and 5 mg of RNA from cash sample was translated using the wheat germ cell free system in a total volume of 100 m1.
*Control RNA samples used in the different experiments were isolated from hypocotyls without IBA pretreatment, but kept in Hoagland's medium for different periods of time.
The radioactivity profiles, after SDS-polyacrylamide gel electrophoresis (SDS-AGE), of the translated products obtained in the wheat germ cell-free system programmed with poly (A containing RNA from control and auxin-pretreated hypocotyls are depicted in Fig. 2. There are some qualitative and quantitative differences in the radioactivity pco6les obtained in the two cases. In particular, a striking increase in the labeling of the M, 55 000-58 000 protein among the translation products after IBA pretreatment is evident.
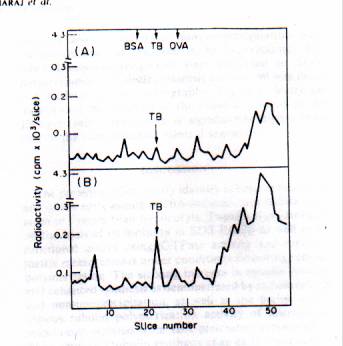
Fig 2. Radioactivity profile of the cell-free translation products programmed with poly (A)-containing RNA isolated from control and IBA-pretreated hypocotyls of Phaseolus vulgaris.
It was suspected that the Mr. 55000-58000 product could be tubulin on the basis of its mobility in SDS polyacrylamide gels and in view of the inhibitory effects of cytochalasin B and colchicine treatment on auxin-induced root formation, implicating microtubule involvement. Therefore, attempts were made to determine whether the concentration of' the Mr. 55000-58000 protein showed
any change in vivo after auxin pretreatment of the hypocotyls segments, From the SDS-polyacrylamide gel analysis of the micros mat protein fraction of the hypocotyls tissues (Fig. 3), it can be seen that there is a striking increase in the content of the Mr. 55 000 protein as early as 18 hr after IBA pretreatment. It is known that tubulin has two distinct subunits, a and b, and the change in the b-subunit (Mr. 58000) concentration is perhaps masked by another protein with the same mobility.
To substantiate further that tubulins were synthesized in response to IBA pretreatment, functional assays were carried out. Tubulin polymerization in a polymerization buffer can be measured using the turbidometric assay procedure and the initiation of polymerization is accompanied by GTP breakdown (9, 10). Tubulin polymerization was thus measured in the presence of microsomal membranes from control and IBA-pretreated hypocotyls, exogenously added tubulin, GTP and other components of the polymerization buffer. Several interesting features are apparent from the results presented in Table 2. The endogenous GTPase activity as well as the tubulin polymerization measured turbidometrically of IBA pretreated hypocotyls membrane in the tubulin polymerization buffer is significantly higher than those of the control membranes. Exogenous addition of tubulin enhances polymerization strikingly in the case of control membranes. Addition of IBA in vitro stimulates tubulin polymerization significantly. The maximum level of polymerization was observed in the presence of exogenous
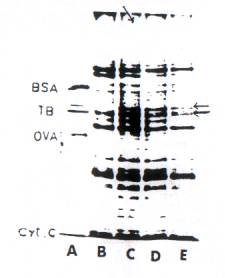
Fig. 3. SDS-Polyacrylamide slab gel electrophoresis of microsomal proteins from control and IBA-pretreated hypocotyls of Phaseolus vulgaris. 100 mg of microsomal protein was analyzed on 8 %, SDS-polyacrylamide slab gels and stained with Coomassie blue. (A) Marker proteins; (B) control (hypocotyls incubated for 12 hr but without IBA pretreatment (C) IBA pretreated, 12 hr; (D) IBA-pretreated, 24 hr; (E) IBA pretreatment, 36 hr. BSA, Bovine Serum Albumin; TB, Tubulin; OVA, Ovalbumin.
Tubulin is the same in control and IBA-pretreated hypocotyl membranes, with the endogenous tubulin polymerization activity of the latter being significantly higher than that of the control membranes.
Finally, tubulin synthesis was measured in vivo by labeling the hypocotyls segments for 6 hr (at the I8 to 24th hr) with 14C-labelled Chlorella protein hydrolysate. The microsomal membrane fraction was brought into
solution and subjected to immunoprecipitation with tubulin antibody as described in the Experimental. The labeled immunoprecipitates were subjacent to SDS polyacrylamide gel electrophoresis and the gel was dried and analyzed by autoradiography. Figure 4 clearly indicates that the labeling of the tubulin subunits in the IBA-pretreated hypocotyls is significantly higher than that of the corresponding control segments.
DISCUSSION
The present studies clearly identify tubulin synthesis as one of the early events in IBA-induced root redifferentiation in French bean hypocotyls. Tubulin was identified on the basis of its mobility in SDS-PAGE as well as by functional assays using GTPase activity and turbidometric measurements under conditions favoring tubulin polymerization. The striking increase in tubulin content and enhanced synthesis as demonstrated by radio labeling and immunoprecipitation, as well as the higher endogenous tubulin polymerization activity of microsomal membranes isolated from IBA-pretreated hypocotyls at 24 hr, implicate tubulin synthesis as an early event in root redifferentiation. It is evident that at least part of the newly synthesized tubulin is still present in an unassembled form (as a monomer pool) at 24 hr after 1BA pretreatment, and perhaps microtubule assembly and reorientation leading to new root formation takes place over a period of 36-40hr after IBA pretreatment. This suggestion is made on the basis that addition of colchicum or cytochalasin B fails to inhibit root production when added after this time.
It can be seen that in the case of IBA-pretreated hypocotyls, radiolabel ling followed by immunoprecipitation and SDS-PAGE (Fig. 4) reveals certain higher M, and lower M, proteins in addition to tubulins, being labeled more than those of the corresponding control tissue, although the total amount of radioactivity taken for immunoprecipitation is the same in both the cases. While there could be non-speoi6c precipitation during the direct immunoprecipitation procedure employed, it is likely that these proteins are tubulin-associated proteins such as 'maps' and other proteins, which are involved in the polymerization process [11].
![]() Table
2. Effect IBA on tubulin polymerization
Table
2. Effect IBA on tubulin polymerization
GTP hydrolyzed Tubulin
![]() Treatment (mol
phosphorus /mg polymerized Protein
per 60 min A at 350nm
Treatment (mol
phosphorus /mg polymerized Protein
per 60 min A at 350nm
Control
Microsomes 0.66 0.02
Microsomes + plant tubulin 1.30 0.72
Microsomes + IBA (in vitro) 0.72 0.04
Microsomes + plant tubulin + IBA 3.30 1.56
(in vitro) IBA – pretreated
Microsomes 1.12 0.13
Microsomes + tubulin 2.01 0.83
Microsomes + IBA (in vitro) 1.86 0.26
Microsomes + plant tubulin + IBA 3.12 1.54
(in vitro)
![]()
Microsomal membranes were isolated from control and IBA – pretreated (24 hr) hypocotyls. Tubulin polymerization was followed by measuring the attendant GTP hydrolysis as well as the change in A at 350 nm during the polymerization reaction. When added in vitro, IBA was used at a concentration of 10 ng / 500ul. The above experimental details are given in the text.

Fig. 4. IBA-induced synthesis of tubulin in vivo. The control and IBA-pretreated hypocotyls were labeled for 6 hr (18-24 hr) with 25 mCi of 14C-labeled Chlorella protein hydrolysate. The microsomal fraction providing 5 x 10-6 cpm of trichloroacetic acid perceptible radioactivity was taken for immunoprecipitation in both cases. (A) Control; (B) IBA-pretreated. TB, Tubulins (a and b units). The tubulin bands amounted for 0.02 and 0.14% of total microsomal proteins labeled in the case of control and IBA pretreatment, respectively.
Another significant observation is the effect of IBA in promoting tubulin polymerization when added in vitro) (Table 2). It appears that the hormone enhances tubulin synthesis as well as having a direct effect on the polymerization process. The effect of IBA on tubulin synthesis is perhaps a transcriptional effect, as indicated by the enhanced amount of Mr., 55 000- 58 000 protein formed in the wheat germ cell-free system programmed with RNA isolated from IBA pre-treated hypocotyls. In addition, the hormone treatment enhances the total messenger RNA activity significantly and at least some of the proteins coded could be tubulin-associated proteins governing the polymerization and orientation of the microtubule assembly, leading to redifferentiation and adventitious root formation.
EXPERIMENTAL
Seeds of Phaseolus vulgaris L. were purchased from the Lalbagh Seed Nursery. Bangalore. India. 14C-Labelled Chlorella protein hydrolysate (27 Ci/m atom C) was purchased from Bhaba Atomic Research Centre; Bombay [3H] Leucine (105 Ci/mmol) was purchase from the Radiochemical Centre,
Amersham. U.K. Oligo dT-cellulose was purchased from Collaborative Research Inc., Waltham, and U.S.A. All other biochemicals were purchased from Sigma.
Effect of colchicine, cytochalasin B and actinomycin D on root production. The conditions for root production in the hypocotyls segments of Phaseolus vulgaris have been described in ref. (8). Briefly, the hypocotyl segments (8-10 cm) obtained from the seedlings were washed thoroughly and then placed in half strength Hoagland's nutrient tedious. Hormone treatment involved the addition of IBA to the medium at a conc. of 10 mg/ml and the hypocotyls were exposed for 30 min. They were then washed thoroughly, placed in fresh medium without the hormone. And the number emerging roots was counted after 72 hr. To prevent bacterial contamination. Ampicillin (30 mg/ml) was added. The effects of Actinomycin D (l0 mg (ml), cytochalasin B (10ug/ml, and colchicines (2.5mg/ml) on root production were tested by adding these components to IBA pretreated hypocotyls after different periods. The hypocotyls were exposed to these compounds for 6hrs, washed thoroughly, and then transferred to fresh media without the inhibitors. The number of emerging roots in each case was counted at the end of 72 hrs.
Quantitation of translatable messenger activity: For this purpose, total and poly(A-containing RNAs were isolated from control and IBA pretreated hypocotyl segments using phenol-CHCl3 extraction procedure [12] Poly(A)-containing RNA was isolated from total RNA as described in ref[13].
The messenger activity of the RNA preparation was assayed in the wheat germ cell-free system [14]. The assay mixture in a vol. of 100 ul contained Hepes, 20 mM; GTP, 20 mM; ATP 1 mM; Phospho creatine, 8 mM; Mg(OAc)2, 3 mM; KCI, 100 mM; DTT, 2 mM; amino acid mixture without Leucine, 25 mM each; (105 Ci/mmol), 20 uCi; and S-30 fraction, 1 A:60 unit of RNA (an optimum concn. in the range giving a linear response). The mixture was incubated at 25° for 60 min and 5 ul portions were transferred to Whatman filter paper discs. Washed with hot and cold CCI3COOH (TCA) and then with Et2O twice, and used for measuring radioactivity incorporated into total proteins. The radioactivity incorporated into the TCA perceptible protein product at identical concentrations of different RNA preparations is taken as the measure of messenger activity.
Radioactivity profile of the cell free products: The labeled cell free products were precipitated with cold TCA (10 % w/v in 6na1 concn) and washed with Et2O. The pellet was dissipated with 2 % (w/v) SDS containing 10 mM Tris-HCI (pH 6.8), 5 % b mercaptoethanol and 10% (v/v) glycerol at 100° for 2 min. The protein products were analyzed using 8 % SDS-polyacrylamide slab gels [15].The gels were stained with Coomassie blue, distained, and then each slot was cut into 1.5 mm slices. The slices were digested with 0.5 ml H2O2 (30% soln) at 60° overnight and the radioactivity was measured using 0.5 % (w/v) 2, 5-diphenyloxazole in Triton-toluene (1:2, v/v) as solvent.
Analysis of membrane proteins: For this purpose, IBA pretreated and control hypocotyl segments were homogenized in 0.25 M sucrose containing 20 mM KPi buffer, pH 7.2 and 1 mM phenylmethylsulphonyl fluoride. The homogenate was filtered through 2 layers of cheesecloth and the filtrate was centrifuged at 10 000 g for 15 min. The supernatant was spun at 105 000 g for 90 min and the pellet was used as the microsomal fraction. The proteins were analyzed using 8% SDS-polyacrylamide slab gels.
Tubulin polymerization: It was followed by assaying two parameters. In one case, the attendant GTPase activity was measured in the presence of microtubule reassembly buffer [9]. The reassembly buffer in a total vol. of 500 ml contained 5 mM morpholine ethane sulphonate (MES) buffer (pH 6.8), 1mM, EGTA. 0.5 mM MgCI2 50 mM KCI, 100 mg of microsomal protein fraction. And 20 mg of tubulin obtained from either rat brain or hypocotyl segments. After the reaction mixture was equilibrated in a shaking water bath at 27% for 10 min, the reaction was initiated by the addition of GTP (1 mM in final concn). The reaction was terminated at different time intervals by adding 50% TCA to a final concn of 10%. The tubes were kept at 00 C for 30 min, centrifuged, and the supernatant was used for the estimation of Pi content by the method of ref. [16].
In the other method, tubulin polymerization was followed using the turbidometric assay procedure [17]. Phosphocellulose purified plant tubulin (1 mg/ml) was incubated in MES buffer (100 mM MES, 0.5 mM MeC12=, 1 mM EGTA and 50 mM KCI, pH 6.8) in the presence of microsomal membranes from control and IBA-pretreated hypocotyls at 37°. The reaction was initiated by the addition of GTP and A was measured at 350 mm as a function of time.
Synthesis of tubulin in vivo: After 18 hr of incubation in half strength Hoagland's medium, both control and IBA-pretreated hypocotyl segments (2-4 cm length) were placed in 5 ml of fresh medium containing 25 mCi of 14C-Iabeled Chlorella protein hydrolysate. After another 6 hr, the segments were thoroughly washed and the microsomal fraction was isolated. The membrane proteins were rendered soluble in 20 mM Tris-HCI (pH 7.4) containing 0.15 M NaCl, 1% Na deoxycholate and 1 % Triton X-100. Immunoprecipitation of tubulin was carried out by the addition of a 2-fold excess of anti-tubulin I8G based on the equivalence point. The incubation was carried out at 37° for 30 min and then at 4° overnight. The ppts were washed with the immunoprecipitation buffer and finally with Tris-NaCl buffer. The immunoprecipitate was then analyzed on SDS-polyacrylamide gels (8%) as described earlier and subjected to fluorography [18]. The tubulin bands were also sliced digested with H2O2 and the radioactivity was measured.
A B
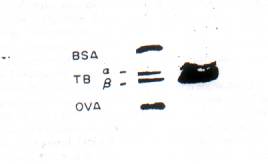
Fig. 5. Purification of tubulin from the hypocotyls of Phaseolus vulgaris. Experimental details are given in the text. (A) Marker protein: BSA, bovine serum albumin: TB, tubulins (a and b units): OVA, Ovalbumin. (B) Purified plant tubulin (3o mg protein).
Other procedures: Antibodies were raised in rabbits for rat brain tubulin prepared according to the method of ref. [19], with certain minor modifications. The IgG fraction was prepared from the antisera and its cross-reaction with purified plant tubulin and the plant microsomal membrane fraction was ascertained by the Ouchterlony technique. Plant tubulin was isolated from IBA pretreated hypocotyl segments by the procedure employed for the isolation of rat brain tubulin. An SDS-polyacrylamide slab gel Electrophoretic profile of the final preparation is given in Fig.5. The anti-tubulin IgG was titrated against the plant tubulin. I8G was also titrated against the plant microsomal membrane proteins rendered soluble by procedures described above and at the equivalent point, 500 mg 1gG proteins was found to immunoprecipitate 3.2 mg of the microsomal proteins quantitatively. Protein content was measured by the method of ref. [20].
Acknowledgements--G.R.K. gratefully acknowledges the help and encouragement given by the University Grant Commission, New Delhi and the authorities of the National College, Bangalore.
REFERENCES
1. Van Overbeck. J. (1961) in Encyclopedia of Plant Physiology (Ruhland, H., Ed.). Vol. XIV, p. 1145. Springer Berlin.
2. Trewavas, A.J. [1968) Arch. Biochem. Biophys. 123, 324.
3. Datko, A. H. and Maclachlan, G. A. (1968) Plant Physiol. 43, 735.
4. Trewavas, A.J. (1976) in Molecular Aspects of Gene Expression in Plants (Bryant, J. A., ed.), p. 249. Academic Press, New York.
5. Davies, E. and Larkins, B. A. (1973) Plant Physiol. 52, 339.
6. Travis, R. L. and Key, l. L. (1976) Plant. Physiol. 57, 936.
7. Travis, R. L., Anderson, J. M. and Key, J. L. (1973) Plant Physiol. 51, 608.
8. Kantharaj, G. R., Mahadevan, S. and Padmanabhan, G. (1979) Phytochemistry 18, 383.
9. Jacobs, M., Smith, H. and Taylor, E. N. (1974) J. Mol. Biol. 89, 455.
10. Timashoff, S. N. and Grihann, L. M. (1980) Annul. Rev. Biochem. 49, 565.
11. Krischner. M. W. (1980) Int. Rev. Cytol. 54, I.
12. Padmanabhan. G., Hendler, F., Patzer, J., Ryan, R. and Rabinowitz, M. (1975) Proc. Natl. Acad. Sci. U.S.A. 72, 4293. 13. Penman, S. (1966) J. Mol. Biol. 17, 117.
14. Roberts, B. E. and Patterson, B. M. (1973) Proc. Natl. Acad. Sci. U.S.A. 70, 233.
1 5. Lamellae, U. K. (1970) Nature (London) 227, 680.
16. Fiske. C. M. and Subba Rao, Y. (1925) J. Biol. Chem. 66. 375.
17. Gaskin, F., Cantor, C. R. and Shelanski, M. L. (1974) J. Mol. Biol. 89, 737.
18. Bonner, W. M. and Laskey, R. A. (1974) Euro. J. Biochem. 46. g).
19. Weingarten, M. D., Littman, D., Suter, M. and Kirschner, M. W. (1974) Biochemistry 13. 5529.
20. Lowry, O.H. Rosenbrough, N. J., Farr, A. L. and Randall. R. J (1957) J. Biol. Chem. 193. 265.