Paper 9
Indole Butyric Acid Induced Early Molecular Events
During Adventitious Root Formation.
G. R. KANTHARAJ* and G. PADMANABHAN**
* Dept. of Botany, The National Collage, Bangalore – 560 004, India. (Correspondence address)
Dept. of Biochemistry, Indian Institute of Science, Bangalore - 560 012,
India
INTRODUCTION
Adventitious root initiation in higher plants is a phenomenon of morphogenesis controlled and regulated by phytohormones. During auxin induced root initiation, cells in the pericycle region, especially over the vascular bundles undergo redifferentiation and develop into root primordial [190,191]. Anatomical studies of root initiation in pea epicotyls and soybean hypocotyls reveal that parenchymatous cells in the cortical region appear to be swollen after 24ht of auxin treatment. After 48hr, cell in the region of root formation start disintegrating and form lacunae. After 3 days, root primordial become recognizable and eventually they occupy the lacunae [196].
In the scheme of molecular event leading to root formation, an increase in RNA and protein synthesis by several folds over a period of 3 days has been observed [196]. In soybean hypocotyls the increase in RNA synthesis occurs after a lag of 1-3 ht. Further, the labeling studies indicate a preferential incorporation of the label into rRNA and tRNA, as compared to AMP-rich RNAs [72,151]. Along with the increase in RNA synthesis, a shift from monosomes to polysomes has also been detected in auxin treated soybean hypocotyls [77]. The shift is inhibited by actinomycin D, 6-methylpurine and cordycepin, but not by 5FU; thus suggesting an increased mRNA synthesis. Ribosomal activation through the synthesis of specific proteins of 60s ribosomes has also been implicated [166].
Thus, it is of interest to examine the very early effects of auxin treatment (1-3hr) on the rate of synthesis of macromolecules and their impact on the later events leading to root formation in the hypocotyls of Phaseolus vulgaris .L.
MATERIALS AND METHODS
Materials:
Seeds of Phaseolus vulgaris. L. have been procured from Lalbagh seed Nursery center, Bangalore , India. [14C] Chlorella protein hydrolysate (27 Ci/m alpha carbon and [32p] orthophosphate (carrier-free) were purchased from Babha atomic Research center, Bombay, India. [3H]-Leucine (105 Ci/mmole) and [3H]-Uridine (27 Ci/mmole) were purchased from Radiochemical Centre, Amersham, Bucks, U.K. Oligo dT-cellulose was purchased from Collaborative Research Inc., Waltham, Massachusetts, U.S.A. Poly (U)-sepharose was purchased from sigma Chemicals co., St. Louis, U.S.A. All other chemicals used were of Analytical Grade.
Methodology:
Morphological Studies:
Phaseolus vulgaris L. seedlings were raised in sand, in a dark chamber. When the hypocotyls attained a height of 15-20 cm, they were cut at the base. Hypocotyls of 8-10 cm length were washed in tap water for 30 minutes and kept in required volume of half strength Hoagland’s nutrient solution [220], with or without plant hormones. In all long-term experiments a 12 hr day period was maintained using fresh medium everyday. To prevent bacterial contamination, 30 hg /ml of Ampicillin (Hindustan Antibiotics Ltd.) was added to the culture medium. The number of roots produced was counted at the end of 4th day.
For anatomical studies, 1 cm length hypocotyls segments were excised and fixed in Conroy’s solution (70 ml of 70% ethanol, 20 ml chloroform and 10 ml of glacial acetic acid). Then the segments were embedded in paraffin wax. Microtome sections (10 m thickness) were taken and processed as described by Jensen (1962) [230]. Microphotographs were taken using 100 ASA panchromatic films.
Protein synthesis in vivo
Protein synthesis in excised hypocotyls was followed by placing the required amount of tissue (suitably treated and washed) in a known amount of half strength Hoagland’s solution containing the required amount of [14 C] – chlorella protein hydrolysate. The different treatments given, the amount of tissue and the label used are described in the legends of respective tables and figures.
The labeled segments were thoroughly washed with distilled water to remove any adhering radioactivity and the tissue homogenate was prepared by grinding in O.o2M sodium phosphate buffer (pH 7.0) in the cold (0-4o). Perchloric acid was added to the homogenate to a final concentration of 0.3M (v/v) and the precipitate recovered by centrifugation after standing for a few hours in cold. The protein was extracted from the precipitate in 1N NaOH, neutralized and reprecipitated by adding 50% trichloroacetic acid (TCA) to a final concentration of 10% (v/v). The precipitate was successively washed with 10% hot TCA (90o C), 10% cold TCA, ethanol-ether (3:1, v/v) and finally with ether. The final preparation, after the removal of ether was dissolved in formic acid and aliquots were planchetted on Whatmann No.3 filter paper discs, dried and used for radioactivity measurements. Similar aliquots, after neutralization, were also used for protein estimation by the method of Lowry et al [221].
The specific radioactivity of the amino acid pool was estimated using the 0.3M perchloric acid supernatant. The supernatant was neutralized and aliquots were used for radioactivity measurements. Total amino acid content was estimated by the method of Rosen [222].
Protein synthesis in vitro
The procedure used for the isolation of polysomes from the hypocotyls segments were as described by Anderson et al. [223]. The tissue was homogenized in 5 volumes of buffer a (containing 50 mM Tris-HC1 (pH 7.5), 5 mM MgC12 , 15 mM KCL, 0.6 mM DTT and 300 mM sucrose). The homogenate was centrifuged at 20000xg for 15 min and 2 ml of the supernatant was layered over 3ml of 1.5M sucrose pad and centrifuged at 150000xg for 2 hr. The pellet was suspended in the homogenizing buffer A. Another batch of homogenate was centrifuged at 20000xg for 15 min and the supernatant was centrifuged at 150000xg for 2 hr directly without layering on sucrose. The final supernatant was passed through a G-25 sephadex column (20xl.5cm) which was already equilibrated with homogenizing buffer. Fractions with maximum A280 values were pooled and used as the source of soluble enzymes (8-150 supernatant fractions).
The in vitro protein synthesizing incubation mixture in a total volume of 100 m1 contained 20mm HEPES buffer (pH 7.5), 5 mM MgCl2, 50 mM KCL, 0.4 mM ATP, 0.12 mM GTP, 5 mM phosphoenol pyruvate, 2 enzyme units of pyruvate kinase, 25 mM each of a mixture of amino acid minus leucine, 100 mg RNA from polyribosome, 100mg protein of S-150 supernatant fraction and 1 mCi of [3H] – leucine (105 Ci/mmole). After incubating the mixture for 30 min at 37oC, aliquots (20 m1) were applied on to filter paper discs. After air-drying, the discs were successively washed with hot TCA (10%) and cold TCA (5%), ethanol –ether (3:1) and finally with ether. The discs were then counted for radioactivity.
RNA Synthesis:
The hypocotyls tissue, after hormone treatment, was incubated for 30min in half strength Hoagland’s nutrient medium containing either carrier free [32 P] – orthophosphate or [3H] –uridine. After incubation the tissue was washed thoroughly with water, and then total RNA was extracted by phenol-chloroform procedure [224] with minor modifications.
Isolation of total RNA:
In these experiments, in order to eliminate RNase contamination, all glass wares were washed in acid and baked overnight in hot air oven (200oC). All aqueous solutions were autoclaved.
The required amount of hypocotyls tissue was homogenized in equal volumes of 0.1M Tris-acetate (pH 9.2) buffer (containing 1% SDS, 0.001M EDTA, 0.1% diethylpyrocarbonate) in a pre-cooled all-glass mortar and pestle. To the homogenate equal volumes of phenol (saturate with buffer) – chloroform (1:1, v/v) was added. After constant stirring for 20-30 min, the extract was centrifuged at 10000kg for 10 min. The aqueous phase was carefully transferred to a separate flask. The interphase was repeatedly extracted with the addition of homogenizing buffer. The aqueous phases were all pooled and equal volumes of phenol-chloroform was added and centrifuged at 10,000kg for 10 min. The aqueous phase was removed and sodium chloride was added to a final concentration of 0.15M. RNA was precipitated with the addition of 2.5 volumes of ethanol and the mixture was left overnight at – 20oC. The pellet was collected by centrifugation at 15,000 kg for 10 min. The total RNA thus obtained was washed successively with 2 volumes of 70% ethanol containing 2M potassium acetate and thrice with 3M sodium acetate (pH 6.8) and finally reprecipitated with ethanol.
Isolation of poly (A) – containing RNA:
Poly (A)-containing RNA was isolated by the methods of Padmanabhan et al. (1975) [225]. Total RNA was suspended is 0.01M Tris-HC1 (pH 7.5) buffer containing 0.5M sodium chloride and 0.5% SDS (high-salt buffer). About 250 A 260 units of total RNA was loaded on to 2.5 ml of poly (U) – Sepharose (0.5 gms) or Oligo dT-cellulose (0.5 gm) column equilibrated with high – salt buffer. The flow rate was adjusted to 1 ml/min. The column was first washed with 10ml of high – salt buffer without SDS. Poly (A) containing RNA was eluted with o.o1M Tris-HC1 (pH 7.5) buffer. The poly (A) – RNA thus obtained was subjected to another cycle of affinity chromatography. To the eluate potassium acetate was added to 0.15M concentrations and poly (A)–RNA was precipitated with the addition of 2.5 vol of ethanol. After keeping the RNA mixture for 16-20 hrs at – 20oC, Poly (A)-RNA was collected by centrifuging at 27000xg for 15 min.
Determination of specific activity of RNA:
In some experiments labeled precursor incorporation into RNA was also followed by measuring the radioactivity released after 0.3M KOH hydrolysis of the 0.3M perchloric acid precipitate obtained with the homogenate. For measuring specific radioactivity of the free nucleotide pool, the supernatant obtained after precipitation of the homogenate with 0.3M HCl04 was treated with activated charcoal. The charcoal pellet was washed repeatedly with water and finally the nucleotides were eluted with 50% ethanol containing 0.3M NH4OH [226]. Aliquots were used for measuring A260 as well as radioactivity.
Analysis of the buffer-extractable proteins labeled in vivo and in vitroPreparation of the buffer-extractable protein fraction labeled in vivo:
Hormone treated hypocotyls tissue (1 gm) was labeled by incubating in 2 ml of Hoagland’s nutrients medium containing 25 mCi of [14 C] – chlorella protein hydrolysate for 30 min. Then the tissue was homogenized in a buffer containing 0.02M sodium phosphate (pH 7.0), 0.25M sucrose and 10mM PMSF. The homogenate was centrifuged at 10,000 rpm for 10 minutes. The supernatant proteins (250 mg) were analysed by SDS-gel electrophoresis and the radioactivity profile of the separated proteins determined.
Preparation of the protein fraction labeled in vitro:
The labeled proteins synthesized in the homologous cell-free system described were precipitated with 10% TCA. After standing for few hours, the precipitate was collected by centrifugation and washed successively with 10% cold TCA, ethanol-ether (v/v, 3:1) and finally with ether. The dried pellet was used for SDS –gel analysis and measurement of radioactivity profile of the separated proteins.
SDS-Gel electrophoresis:
SDS-polyacrylamide slab gel (8%) electrophoresis was carried out according to Lammli’s procedure [227]. Tris-glycine (pH 8.8) buffer containing 0.1% SDS was used as the electrophoresis buffer. The protein samples were taken in the dissociating buffer containing 10 mM Tris-HC1 (pH 6.8), 10% glycerol, 2% SDS, 5% (v/v) b-mercaptoethanol and heated in boiling water (100oC) for 2 minutes. The protein samples were loaded into the slots and electrophoresis was carried out at 20 mA constant current (0-4oC) till the marker dye bromophenol blue reached the bottom of the gel. The gels were removed and fixed in a solution containing 10% acetic acid and 40% methanol for 30min, stained with 0.5 %( w/v) Coomassie brilliant blue-R (Sigma) overnight and destained with the same fixative solution. After destaining, the gels were photographed using red filter. The gels were cut into 1mm slices and digested with H2 O2 (30%) at 600C for the purpose of radioactivity measurements.
Radioactivity Measurements:
The H2O2 digested gel slices were counted using Triton-toluene (1:2, v/v) containing 0.5% PPO in a Beckman LS-100 scintillating counter. Filter discs were counted using 0.5% PPO in toluene as the scintillant. Under the conditions used 3H, 14C and 32P were counted with efficiencies of 6%, 35% and 80% respectively.
RESULTS
BASIC PARAMETERS
Certain basic parameters influencing root initiation in the excised hypocotyls sere first determined. The results presented in Table 1 Indicate that the number of roots produced by IBA is proportional to the hormone concentration up to 10 m g/ml. A further increase in the concentration does not significantly increase the number of roots produced. Similarly, 8-10 minutes of treatment with the hormone appears to be
Table 1: Effect of IBA concentration and duration of treatment on root production in the hypocotyl segment of Phaseolus vulgaris L.
|
IBA concentrationm g/ml |
No. Of roots formed/segment (average of 10 segments) |
Duration of IBA treatment (min) |
No. Of roots formed/segment (average of 10 segments) |
|
0 |
1.2 ± 0.6 |
2 |
19.1 ± 1.0 |
|
2 |
45.2 ± 1.3 |
4 |
42.7 ± 2.5 |
|
4 |
58.3 ±1.8 |
6 |
69.7 ± 6.5 |
|
6 |
71.1 ± 1.7 |
8 |
76.6 ± 4.0 |
|
8 |
86.9 ± 1.4 |
10 |
77.9 ± 1.7 |
|
10 |
82.8 ± 1.4 |
12 |
73.8 ± 2.6 |
|
12 |
77.6 ± 1.6 |
20 |
76.6 ± 1.5 |
|
20 |
75.8 ± 1.5 |
30 |
73.6 ± 3.3 |
|
30 |
77.9 ± 1.3 |
|
|
Hypocotyl segments (10 segments of 8-10 cms length each) were placed erect in a flask containing 100 ml of half strength Hoagland’s medium (basal medium). IBA was added to the basal medium in different flasks in increasing concentrations and the effect of IBA concentration on the number of roots formed was examined when the tissues were exposed to the hormone for 30min and then placed in a fresh medium. The effect of the duration of the hormone treatment was examined at an IBA concentration of 10mg/ml. The number of roots formed 72 hr after transfer of the treated hypocotyls to fresh basal medium was counted ± standard error. *Mean means sufficient for maximal root induction. It is also possible to induce root formation in segments of just 2cm length (placed either upright or upside down) and in segments, which are vertically split into four strips (data not given).
ANATOMICAL STUDIES
Several cells of the pericycle, situated over the split exarch xylem strands are enlarged and show few divisions, within 24 hours of auxin treatment. Many of these cells possess large nuclei with conspicuous nucleoli. At the end of the next 24 hours these cells by repeated cell divisions have organized into root primordial. Some pith parenchymatous cells at this time-point start disintegrating and many cortical parenchymatous cells situated around the root primordials are greatly enlarged. By 72 hours, the root primordials push through the cortex and emerge at the surface of the hypocotyls. In the path of the root primordials many cortical cells are found to be either crushed or disintegrated. However, a large central lacuna is found in the pith region. An invariant four row pattern of adventitious root formation is maintained from the base to the tip of the hypocotyls (Fig. 4).
IBA INDUCED RATE OF PROTEIN SYNTHESIS
In order to understand the IBA mediated very early macromolecular events, the rate of protein synthesis
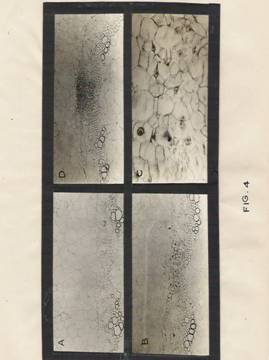
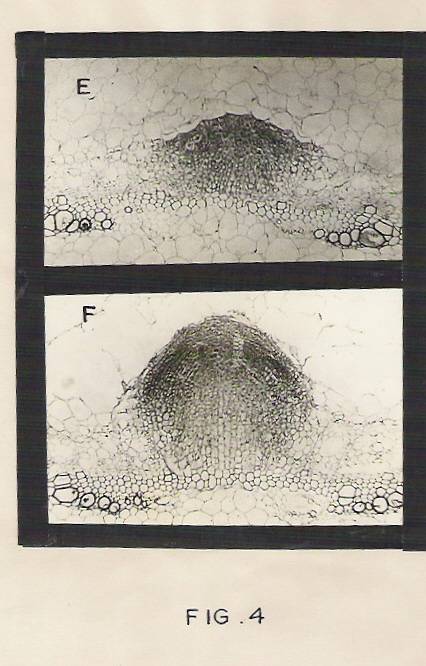
Fig.4: Photographs showing anatomical changes during adventitious root initiation.
a) Transverse section of control hypocotyls segments showing pericyclic cells split exarch xylem strands, endodermis, cortical cells and pith cells. Magnification: X200.
b) Transverse section of hypocotyl segment treated with IBA for 24 hr, showing enlarged pericycle cells above the split exarch xylem strands. Magnification: X 200.
c) A part of Fig. B is magnified to show enlarged nucleoli, nuclei and cells. Magnification: X 800.
d) Transverse section of the hypocotyls treated with IBA for 36 hr showing cell divisions in the pericyclic region. Magnification: X 200.
e) Transverse section of hypocotyls segment. 48hr after IBA treatment showing young root primordium, Magnification: X 200.
f) Transverse section of hypocotyls segment, 72 hr after IBA treatment showing well developed adventitious root penetrating through the cortex. Enlarged and disintegrating cells are also seen in the cortex. Magnification: X 200.
Was determined at different time-intervals. The results given in table 2 indicate that IBA enhances the rate of protein synthesis two fold as early as 30 minutes of treatment. To determine, whether the early increase in protein synthesis is mediated by increased RNA synthesis, the effect of Actinomycin D on protein synthesis was studied and the results are presented in Table 3. It is clear from the table that the early increase in protein synthesis in response to IBA is insensitive to actinomycin D. However, the increase in protein synthesis 2 hours after IBA treatment is blocked by actinomycin D, there by suggesting that this increase in protein synthesis is mediated by increased RNA synthesis. In all these experiments, striking changes in the specific radioactivity of the free amino acid pool are not detected. In a separate experiment, actinomycin D has been found to inhibit RNA synthesis by over 90% at both time-points (30 min as well as 2 hrs).
IBA-INDUCED RATE OF RNA SYNTHESIS
To extend these results, the rates of RNA synthesis, in response to IBA treatment, were measured using [32P] orthophosphate or [3H] uridine as the labeled precursors. The results (tables 4 and 5) again suggest that IBA pretreatment has on effect on total RNA or poly (A) –containing RNA synthesis at the 30 min time period. However, after 2hts, there is a striking increase in the rate of total RNA as well as poly (A)-RNA synthesis.
ACTIVATION OF PROTEIN SYNTHESIS IN VITRO
It is of interest to examine the mechanism to IBA mediated RNA-independent, increase in early protein synthesis. An attempt has been made to determine, whether the enhanced rate of protein synthesis detected in vivo in hypocotyls segments treated with IBA for 30 min can also be demonstrated in vitro. For this purpose, polyribosome and post-ribosomal supernatant fractions (S-150) were prepared from IBA-treated and control hypocotyls. Protein synthesis was monitored in vitro, using the polyribosomes and S-150 fractions as described in materials and methods. The reaction rate is linear up to 30 minutes. The rate of protein ;synthesis is maximum with the combination of polyribosomes and S-150 fraction prepared from the IBA-treated hypocotyls (Table6) However, the addition of S-150 fraction from IBA treated hypocotyls to control ribosome’s also results in a significant stimulation of protein synthesis in vitro. Further, the addition of IBA (10 ng) to the in vitro protein synthesizing reaction mixtures (100 ml) containing polyribosomes and S-150 fraction from control hypocotyls also results in a measurable increase in the rate of protein synthesis.
Table 2: Effect of IBA on protein synthesis in the Hypocotyl tissue of P.vulgaris
---------------------------------------------------------------------------------------------------------------------Treatment 0 min 30 min 120min
Cpm/ug cpm/ug cpm/ug
---------------------------------------------------------------------------------------------------
(A) (B) (A/B) (A) (B) (A/B) (A) (B) (A/B)
---------------------------------------------------------------------------------------------------
Control 53.6 7.8 0.14 49.6 7.2 0.14 56.0 8.4 0.15
IBA 36.0 11.8 032 45.2 13.2 0.29 43.2 11.8 0.27
--------------------------------------------------------------------------------------------
Hypocotyl tissue (2 g) was placed in 5 ml of basal medium with or without IBA (30 mg/ml), After 30 minutes of exposure, protein synthesis was followed immediately (o min) and at different time intervals after transfer to the fresh nutrient medium using [14 C] – chlorella protein hydrolysate (10 mCi/5 ml). The labeling period was 30 minutes.
Table 3: Effect of IBA on RNA synthesis in the hypocotyl tissue of Phaseolus vulgaris L.
Treatment 0 min 120min
Amino Protein A/B Amino protein A/B
Acid pool Acid pool
A (cpm/ug) B (cpm/ug) A/B A (cpm/ug) B (cpm/ug) A/B
Control
(-) Actinomycin-D 41.6 4.6 0.10 52.4 5.2 0.09
(+) Actinomycin-D 45.4 3.8 0.08 46.4 4.8 0.10
IBA
(-) Actinomycin-D 39.6 6.8 0.17 42.8 9.6 0.22
(+) Actinomycin-D 37.0 7.2 0.19 48.4 4.2 0.08
--------------------------------------------------------------------------------------------
Hypocotyl tissue was treated with Actinomycin-D (100ug/ml /2g) for15 min before the addition of IBA (30ug/ml). After 30 min, the tissue was washed thoroughly and placed in a fresh basal medium. Protein synthesis was followed immediately (0 min) and 120 min after transfer to the fresh medium using [14C]-chlorella hydrolysate 910uCi/5ml). The labeling period was 30 min.
Table 4: Effect of Actinomycin-D on IBA-mediated increase in the rate of protein synthesis in the hypocotyls tissue of Phaseolus vulgaris L.
|
Treatment |
0min |
0 min |
0 min |
120 min |
120 min |
12 min |
|
|
Ntds pool Cpm/A260 |
Total RNA Cpm/A260 |
Poly-A RNA Cpm/A260 |
Ntds pool Cpm/A260 |
Total RNA Cpm/A260 |
Poly-A RNA Cpm/A260 |
Control |
22470 |
2420 |
670 |
18500 |
12500 |
1690 |
IBA |
20560 |
2330 |
620 |
19580 |
3500 |
1190 |
|
|
|
|
|
|
|
|
The experimental details are as given in Table 3. [32P]-orthophosphate 9240 Ci/10 ml/10gmtissue) was added immediately (0 min) as well as 120 min after transfer of hormone-treated hypocotyls tissues to fresh basal medium. The labeling period was 30mn. Total RNA as well as poly (A)-containing RNA synthesis was followed as described in the text.
Table 5: Effect of IBA on RNA synthesis in the hypocotyls tissue of P.vulgaris L.
--------------------------------------------------------------------------------------------
Treatment 0 min 120 min
------------------------------------------------------------------------------------------------
Total RNA Poly (A) RNA) Total RNA Poly (A) RNA
(Cpm/A260 (cpm/A 50 260 (cpm/A260 (cpm/50 A260
Unit) units of total RNA) unit) units of total RNA)
Control 143 35 153 42
IBA 159 39 309 96
The experimental details are as given in Table 4 except that RNA synthesis was followed using [3H]-Uridine (500 uCi/5ml/10m tissue).
Table 5: Effect of IBA on RNA synthesis in the hypocotyl tissue
of Phaseolus vulgaris L.
|
Treatment |
O min |
120 min |
||
|
|
Total RNA (cpm/A260 Unit) |
Poly (A)-RNA
(cpm/50 A260 Unit of total RNA) |
Total RNA (cpm/A260 Unit) |
Poly (A)-RNA
(cpm/50 A260 Unit of total RNA) |
|
|
|
|
|
|
|
Control |
143 |
35 |
153 |
42 |
|
|
|
|
|
|
|
IBA |
159 |
39 |
309 |
96 |
The experimental details are as given in Table 4 except that RNA synthesis was followed using [3 H] – uridine (500 mCi/5ml/10 gm tissue).
ANALYSIS OF EARLY PROTEIN PRODUCTS
The above results strongly suggest that I000BA, at very early time periods of treatment, activates the translating machinery. However, to determine, whether this early
Table 6: Effect of Indole Butyric Acid on polysome mediated protein synthesis in vitro.
[3H]-leucine incorporated into TCA perceptible protein (cpm/A 260 units)
Experiment 1 Experiment 2
_______________________________________________________
Control polysome
+ control S-150 21700 15500
IBA polysome
+ IBA S-150 47900 79650
Control polysome
+ control S-150 39400 46100
IBA polysome
+ IBA S-150 24000 22850
Control polysome
+ control S-150 +IBA (10 ng) 29000 33000
Hypocotyl tissue (20gm) was treated with IBA (30 mg/ml), immediately after incubation, the segments were washed in water and polysomes and S-150 supernatant factors were isolated as described in Materials and Methods.
Protein Synthesis in vitro was carried out as described in text. The results obtained in two independent experiments are given.
Increase in protein synthesis is just quantitative or qualitative, labeled protein samples from the in vitro experiments were prepared, as described in the experimental section. Proteins were also labeled in vivo with [14C] chlorella protein hydrolysate for 30 minutes and the buffer extractable proteins were prepared as detailed in materials and Methods section. Protein samples thus obtained were electrophoresed on SDS –polyacrylamide gels. As there was no significant change in the visible protein pattern on the gels, the gels were sliced and counted to determine the radioactivity profiles of the proteins. The radioactivity profiles (Fig. 5 and 6) suggest that the proteins synthesized in vitro and in in vivo in response to IBA treatment are different quantitatively and also qualitatively.
EFFECT OF IBA MEDIATED EARLY INCREASE IN PROTEIN SYNTHESIS ON LATER MOLECULAR EVENTS:
EFFECT ON ROOT FORMATION AND PROTEIN SYNTHESIS:
It is of importance to find out whether the early IBA promoted protein synthesis has any effect on the later transcriptional and translational events leading to root formation. Therefore the segments were treated with cycloheximide and IBA to block early IBA induced protein synthesis. After 30 min, the segments were washed thoroughly to remove the CHI and transferred to fresh Hoagland’s medium.
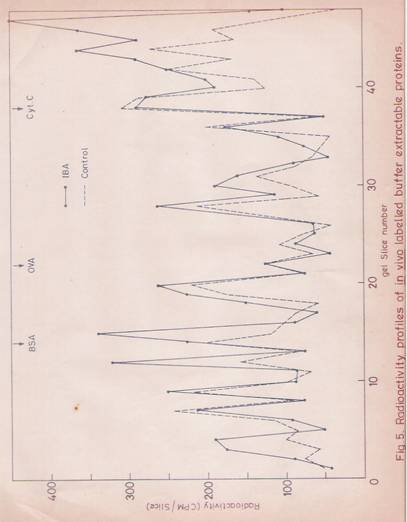
Fig. 5: Radioactivity profiles of in vivo labeled buffer-Extractable Proteins.
Hypocotyl segments of 1 gm (4 cm length) were placed in 2 ml of half strength Hoagland’s medium with or without IBA (30 mg/ml). Immediately 20 mCi of [14C]-Chlorella hydrolysate was added. After incubating for 30 min, hypocotyls were washed thoroughly with water. Protein samples were prepared as described in Materials and Methods. Protein content of 250 mg was loaded onto each slot of the gel. After electrophoresis, the stained gels were sliced and counted for radioactivity.
Control (- - - - - - -- - -)
![]() IBA ( )
IBA ( )
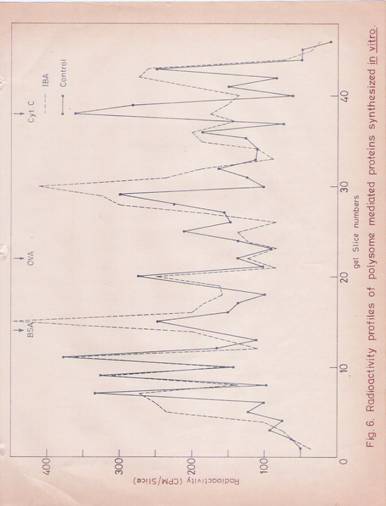
Fig. 6: Radioactive profiles of polysome Mediated Proteins Synthesized in vitro.
The labeled proteins synthesized in vitro were analyzed by SDS-gel electrophoresis. The gels were sliced and counted for radioactivity.
![]() ( ) Proteins
obtained form the reaction mixture containing S-150 supernatant and S-150
polysome fractions from untreated segments.
( ) Proteins
obtained form the reaction mixture containing S-150 supernatant and S-150
polysome fractions from untreated segments.
(- - - - - - - - - - -) Proteins obtained from the reaction mixture containing S-150 supernatant and S-150 polysome fractions obtained from the segments treated with IBA FOR 30 minutes.
To begin with rooting was examined after 72 hr. The results (Table 7) indicate that the inhibition of IBA-induced early protein synthesis has delayed root initiation considerably and the numbers of roots ultimately formed are also decreased by 75%. In this context, it is of relevance to examine how long the inhibitory effect of CHI on protein synthesis persists. For this purpose, the hypocotyls were treated with IBA or IBA + CHI for 30 minutes, then washed thoroughly with water and placed in a fresh basal medium. The rate of protein synthesis was determined immediately as well as at different time-intervals after transferring the hypocotyls to fresh nutrient medium containing [14C]-chlorella protein hydrolysate. The results (table 8) suggest that the inhibitory effect of CHI is significant up to 60 minutes after transfer to the fresh medium. However, no significant inhibition of protein synthesis is observed after 120 minutes of transfer.
EFFECT ON LATER TRANSCRIPTIONAL EVENTS
The effect of inhibition of early IBA-triggered protein synthesis, on the later transcriptional events has also been studied. Hypocotyls were treated with CHI and IBA simultaneously for 30 min and than the tissue were washed thoroughly and transferred to fresh medium. At different time points, the hypocotyls segments wore incubated with Hoagland’s nutrient medium containing [32P]-orthophosphate for 30 minutes.
Table 7: Effect of cycloheximide treatment on root initiation by IBA in the hypocotyls tissue of Phaseolus vulgaris L.
Pretreatment No. of roots formed/segment (Average of 10 segments)*
____________________________________________
72 hr 96 hr 120 hr
________________________________________________________________
Basal medium nil nil nil
IBA 77.6±1.3 79.8±1.4 80.1±5.3
IBA +
Cycloheximide nil nil 21.1±2.4
--------------------------------------------------------------------------------------------
The hypocotyls segments of 8cm length were treated with IBA (30 mg/ml) were added to some flasks. The segments were exposed to IBA ± additives for 30min, washed thoroughly and then transferred to fresh basal medium. The number of roots formed was counted at different time’s intervals.
* Mean ± standard error.
Table 8: Rate of protein synthesis after exposure of the hypocotyls tissues of P.vulgaris L to IBA and Cycloheximide.
|
Treatment |
0 min |
30 min |
120 min |
Cpm/ug cpm/ug cpm/ug
amino acid/protein amino acid/protein amio acid/protein
pool pool pool
(A) (B) (B/A) (A) (B) (B/A) (A) (B) (B/A)
Basal medium 63.6 7.8 0.14 49.6 7.2 0.14 56.0 8.4 0.15
Basal medium+ 36.0 11.8 0.32 45.2 13.2 0.29 43.2 11.8 0.27
IBA
Basal medium+ 52.4 4.6 0.08 46.0 7.6 0.16 44.010.4 0.23
IBA+CHI
--------------------------------------------------------------------------------------------
Hypocotyls tissue (2gm) was placed n 5 ml of basal medium with or without IBA (30ug/ml). T some flasks containing the IBA-medium, CHI 10ug/ml) was added. The segments were exposed for 30min, washed thoroughly and transferred to a fresh basal medium. Protein synthesis was followed immediately (0 min) and at different intervals after transfer o the fresh basal medium using [14C] - chlorella hydrolysate (10 uCi/5ml). The labeling period as 30 min.
Table 9: Effect of cycloheximide on Indole Butyric Acid induced RNA synthesis in the hypocotyls of Phaseolus vulgaris L.
________________________________________________________________
Treatment Time Nucleotide Total RNA B/A Poly (A)±
Pool RNA
(cpm/A 260) (cpm/A 260) (cpm/30A 260)
A B units
_______________________________________________________
Control 2 Hr 22200 3080 0.13 1530
6 Hr 24500 4000 0.16 1280
12 Hr 23500 3500 0.14 1380
24 Hr 29800 3480 0.11 1460
IBA 2 Hr 19840 3730 0.18 2110
6 Hr 22100 5150 0.24 3110
12 Hr 21500 5800 0.25 4200
24 Hr 28040 7000 0.27 5200
IBA+CHI 2 Hr 23000 2675 0.11 1110
6 Hr 22350 3800 0.17 1410
12 Hr 23100 5400 0.23 3100
24 Hr 30500 6500 0.21 3380
Hypocotyl segments were pretreated with cycloheximide (CHI) (10 mg/ml) for 30 minutes. The IBA (30 mg/ml) was added to the same flask. After incubation for 30 min, segments were thoroughly washed in running water for 5 min and transferred to fresh basal medium. T different time-points, 10gm of segments were removed and incubated for 30 min in 5ml of the basal medium containing 200 mCi of [32P]-orthophosphate. Them the segments were washed in water and processed for measuring the rate of total and poly (A)-RNA synthesis as described in Materials and Methods. The data presented are the mean of two independent experiments.
Then the total, as well as poly (A)-containing RNA were isolated and the rate of RAN synthesis was determined. The results in the TABLE 9 show that the inhibition of IBA mediated early protein synthesis by CHI, has a significant repressive effect on total as well as poly (A)-RNA synthesis up to 6 hrs after treatment. But 12 hrs after the transfer to the fresh basal medium, the rate of RNA synthesis recovers from repression and rises to the levels of IBA-induced rate of RNA synthesis.
DISCUSSION
An absolute requirement of apical buds for root initiation has been suggested [228, 229]. In the present study, the hypocotyls segments obtained after excision of cotyledons, leaves, apical and axillary buds, followed by through washing of the segments in running water, has been found to initiate hardly any roots in the basal medium. Exposure of the hypocotyls tissue to the hormone for a very short period of 10 min is sufficient to induce maximum root formation. These results suggest that the root induction is primarily due to exogenously applied hormones and that perhaps the hormone induces the cells to undergo redifferentiation into root primordial.
It has also been suggested that the induction of roots on stem cuttings by the application of hormone is unusual in showing no apparent or plateau in the dose-response curve, with the higher concentration of auxin eliciting a greater number of roots [196]. In the present system employed, the IBA-induced root formation is found to be dependent on hormone concentration and duration of exposure to the hormone.
Morphological studies show that only pericycle cells over the split, exarch xylem strands show greater enlargement of cells, with few cell division at the end of 24 hr of auxin treatment. The presence of large nucleus with conspicuous nucleolus in these cells indicates the intense biochemical activity in response to IBA in this time period. At the end of 48 hrs, the above cells, by repeated cell divisions, show organization into root primordial. The development of root primordial only in this region and the complete absence of root initiation elsewhere in the hypocotyls suggest that the cells in the above said region have an inbuilt cell potentiality and they alone respond to the applied phytohormone stimulus and develop into new roots. The onset of disintegration of parenchymatous cells in the pith region as early as 24 hrs (leading to the formation of a large central cavity and the disintegration of some cortical cells in the pathway of the out growing root primordial at 72 hrs after IBA treatment, are at variance with the observation, that disintegration in the region of root formation at very early stage provides some stimulus for root initiation [196]. In French bean hypocotyls, however, it appears that the disintegration of cortical cells is rather due to the effect of onward growth of the root primordial than the cause of it.
An early effect of IBA is to cause an increase in the rate of protein synthesis. This effect manifested within 30 min after hormone treatment, appears to be independent of a concomitant increase in RNA synthesis. This is proven by the results(1) IBA has no effect on total or poly (A)-RNA synthesis at this early time interval, (2) the increase in the rate of protein ;synthesis in response to IBA is not sensitive to actinomycin D.
This very early effect of IBA can thus be attributed to the ribosomes at the post-transcriptional/translational sites. The fact that the S-150 fraction (cytosol) from IBA pretreated hypocotyls of addition of in vitro stimulates cell-free protein synthesis, indicates that IBA may activate some soluble factor(S) involved in translation. At the same time, maximal rate of protein synthesis in vitro has been observed with ribosomes and cytosol isolated from IBA- treated hypocotyls. This suggests that IBA may also have an effect at the ribosomal level.
The increase in the rate of protein synthesis observed 2hr after IBA treatment is dependent on a concomitant increase in RNA synthesis. At this time point there is an increase in total as well as poly (A)-containing RAN synthesis. Furthermore, actinomycin D blocks the increase in the rate of
Protein synthesis observed 2 hrs after IBA treatment. This data is in agreement with the results obtained in other hormone-department systems [90].
An analysis of early protein products, on SDS-PAGE, of both in vivo and in vitro labeling studies, clearly demonstrate, that among the proteins synthesized in response to IBA, at least some are qualitatively different from the control.
The involvement of IBA-mediated early protein synthesis on the hormone mediated root initiation has been examined by using CHI to block protein synthesis. Inhibition of early protein synthesis using CHI followed by its removal results in a considerable delay in root initiation, as well as the decrease in the number of roots produced. An interesting feature observed is that when cycloheximide is removed, the rate of protein synthesis reaches the IBA induced levels in about 2 hrs. However, the rate of RNA synthesis remains depressed for a longer period and reaches IBA-induced levels only by 12 hrs after the removal of cycloheximide. The inhibition of poly (A)-RNA synthesis is much more striking than that of total RNA. These results indicate that the IBA-mediated early protein synthesis regulates RNA synthesis. The intriguing feature is that the rate of protein synthesis reaches IBA-induced levels much earlier than that of RNA synthesis. At the same time the results also suggest that the IBA-mediated increase in protein synthesis at 2 hrs is sensitive to actinomycin D (Table 3). It may be that the synthesis of different RNA fractions is not equally sensitive to cycloheximide treatment. Bulk of the messenger RNA fraction in this system appears to be poly (A) – and its synthesis/recovery may not be strikingly affected by cycloheximide. Since the rate of synthesis of only total RNA and poly (A)-RNA have been measured, the effects on the synthesis of specific species of messages may not be apparent.
The above results clearly suggest that some of the early proteins synthesized, in response to IBA (30 min-12) hrs) are vital to the process of differentiation. Thus, IBA may activate protein synthesis as early as 30 min, which may in turn activate RNA synthesis leading to the formation of specific proteins required for the re-differentiation process.
SUMMARY
1. Exogenous supply of auxin is required for adventitious root formation.
2. Hypocotyls treated with 8-10 mg/ml of IBA for 8-10 minutes produce maximum roots.
3. The cells situated in the pericycle region over the split exarch xylem respond to IBA treatment and differentiate into root primordials within 48 hr of treatment.
4. Within 30 minutes of IBA treatment protein synthesis is, enhanced and it is insensitive to actinomycin – D. The later increase in protein synthesis is mediated by the IBA-induced RNA synthesis.
5. The very early increase in protein synthesis is due to the activation of some cytosolic factors and ribosomal machinery.
6. The early proteins synthesized in response to IBA are qualitatively different from control hypocotyls.
7. Inhibition of IBA-mediated early protein synthesis by cycloheximide delays and inhibits root initiation considerably.
8. IBA-induced early proteins have a vital role on RNA synthesis specific for root initiation.