Paper 8
Molecular Aspects of Cytokinin's Stymied Action on Auxin Mediated New Root Formation in the hypocotyls of Phaseolus vulgaris L.
G. R. KANTHARAJ* and G. PADMANABHAN**
* Dept. of Botany, The National Collage, Bangalore – 560 004, India. (Correspondence address)
** Dept. of Biochemistry, Indian Institute of Science, Bangalore - 560 012, India
INTRODUCTION
Elucidation of molecular events that regulate organogenesis either in an in vitro or in an in vivo system is a daunting task fraught with many imponderables. It is deja vu that a set of phytohormones control the growth and development of plant body· It is also fiat accompli that any two growth hormones such as auxin or cytokinin, though having their own specific effects on different target tissues, yet their combined action on the same tissue varies and it is concentration dependent (Millar, 1961). The interplay of different hormones at molecular level in inducing or inhibiting one or the other morphogenic structures such as root or shoot or redifferentiation of differentiated tissues into new organs is both befuddling and contentious.
Among many factors, which regulate adventitious root formation, auxin is of prime importance. During auxin induced new root formation either in epicotyls or hypocotyls and other plant tissues, several biochemical changes including increase in DNA, RNA, protein and even enzyme levels have been reported (Davies and Larkins, 1973; Travis et al., 1973; Travis and Key, 1976)· Whether or not the differential gene activity leading root formation involves "Homeotic" kind of gene complex and compartmentalization, as found in Drosophila species, also exists in plant systems cannot be averred for the lack of specific mutants (Scott, 1985).
In earlier investigations, we have preferred excised segments of 3 - 4 cm length of Phaseolus vulgaris, Linn., to in vitro tissue culture methods, for raison d' etre that the segments in response to 5 x 10-5 M concentration of Indole Butyric Acid (IBA) produce 30 - 40 new roots in about 48 hrs (Kantharaj et al., 1979)· During IBA mediated root initiation, early enhancement (30 min) in the rate of protein synthesis was due to auxin activation of protein synthesizing machinery and not due to early increased synthesis of a total RNA including poly(A)-containing RNA. The increased protein synthesis at later stages was augmented by the enhanced RNA including poly (A)-containing RNA synthesis. Among many changes, in IBA enhanced or induced proteins, the 55 - 58 Kd proteins were found in higher quantities, and the same have been identified as Tubulins (a & b units) by their mobility on the gel and also immunological methods (Kantharaj et al., 1985). The present study is actuated to understand molecular events that regulate the inhibition of IBA mediated new root initiation by Benzy1 Amino Purine (BAP) - a synthetic cytokinin.
METHODS
Materials: French bean seeds were purchased from a local seed nursery. Hypocotyl segments of 3 - 4 cm length from the dark grown seedlings were cut. Then they were abated of auxin by washing in distilled water for 30 mins and made certain that none of the segments produced any new roots on their own. Such segments were incubated for 30 mins in half strength Hoagland's medium (Arnold, 1968) containing 5 x 10-5 M IBA or 4·5 x 10-6 M BAP or both. Then the segments were surface washed and further incubated in nutrient medium for the required durations. Aseptic condition was maintained strictly in all experiments.
14C -labeled Chlorella hydrolysate (270/m atom C) and 32p - carrier free orthophosphate were purchased from Bhaba Atomic research Centre, Bombay. 3H –Leucine (105 Ci/mM was) purchased from Radiochemical Centre, Amersham., U.K., Oligo dT-cellulose and Poly(U)-Sepharose were purchased from Collaborative research Inc., Waltham, U.S.A All other biochemicals were purchased from Sigma, U.S.A
Rate of protein synthesis and RNA synthesis: Hormone pre-treated and control segments at different stages of development were exposed to 14C Chlorella hydrolysate or 3 H - UTP or 32P - orthophosphate for 30 mins; then they were surface washed and processed for determining the protein synthesis by the methods of Rosen, 1957 and Lowery, 1957, and RNA synthesis by the methods of Pennman, 1966 and Padmanabhan et al., 1975·
Quantification of mRNA activity and in vitro protein: synthesis Isolated total RNA or poly (A) +RNA were translated in a cell free system derived from wheat germ using 3H - Leucine as the label by Roberts and Patterson method, 1973.
Protein analysis: Membrane proteins and in vitro radiolabel led poly (A) +RNA translated proteins were subjected to SDS-polyacrylamide gel electrophoresis (SDS-PAGE) by the methods of Laemli, 1970. For measuring radioactivity of proteins in the gels, the gels were first stained with Coomassie brilliant blue and then the required gels were sliced to 1-mm pieces, and digested, and counted for radioactivity.
RESULTS
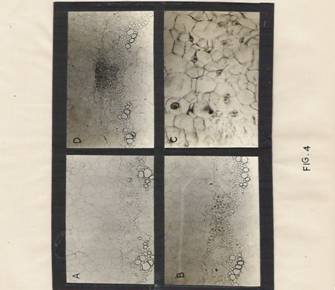
Anatomical and Cytological Changes: The pericyclic cells found in between the split each xylem elements in the hypocotyls treated with auxin were found in transformed state at 12 hr and some of them were mitotically active with periclinal orientation. At 24 hrs, a distinct meristematic dome, for new roots were found in the same locus. On the contrary, in the hypocotyls treated with both IBA and BAP, the same pericyclic cells were in a state of proliferation and no root initials were found. However, the hypocotyls exposed to BAP alone did not show any changes in pericycle region even after 48 hrs, except for the enlarged state of cells (figures 4, A to F)

Rate of protein synthesis: The rate of protein synthesis as measured by the above mentioned method (Table 1) show increased levels in all those segments treated with IBA, BAP or both. Such increased levels of proteins at later stages ware found in all hormone treated tissues.
Table 1: Rate of protein synthesis: Hypocotyl segments of different stages of development were labeled with 14C - Chlorella hydrolysate for 30 mins. From the homogenate, specific activity of proteins (cpm / mg protein = A) and specific activity of total amino acid pol (cpm / mg protein = B) were determined. A / B gives the measure of the rate of protein synthesis.
-------------------------------------------------------------------------------------------- Treatment 30 min 3hrs 6hrs 12 hrs 24hrs
--------------------------------------------------------------------------------------------
Control 0.07 0.08 0.07 0.08 0.11
IBA 0.15 0.19 0.20 0.29 0.46
BAP 0.13 0.14 0.31 0.25 0.40
IBA+BAP 0.11 0.12 0.14 0.30 0.47
--------------------------------------------------------------------------------------------
Table 2.
Rate of RNA synthesis: Hormone treated and control hypocotyls ware labeled for 30 mins with 32 p – Phosphate (Carrier free). Total RNA was precipitation. Specific activity of total RNA i.e., A, in cpm/ A 260 and specific activity of total nucleotide pool i.e., B, in
cpm / A 260 were determined. The rate of RNA synthesis is given by A/B
![]()
![]() TREAMENT 20
mins 2hrs 6 hrs 12hrs 24hrs
TREAMENT 20
mins 2hrs 6 hrs 12hrs 24hrs
Control 0.13 0.12 0.13 0.18 0.18
IBA 0.12 0.20 0.24 0.29 0.33
BAP 0.11 0.12 0.15 0.27 0.28
![]() IBA
– BAP 0.12 0.13 0.19 0.30 0.41
IBA
– BAP 0.12 0.13 0.19 0.30 0.41
Table 3
Quantification of poly (A) +RNA Total RNA was isolated from different hypocotyls segments and then 20 ug of RNA was translated in a cell free system derived from wheat germ containing 3H – Leucine as label. Three separate experiments were conduct and there aliquots for each experiment were counted, and the average cpm for each experiments is presented as a measured as a measure of template activity.
![]()
![]() TREAMENT 12
hrs 24 hrs
TREAMENT 12
hrs 24 hrs
Expt I Expt II Expt III Expt I Expt II Expt III
Control 46640 49500 48000 45750 54300 52300
IBA 75780 80120 78650 136170 142000 138200
BAP 72500 70150 70100 160580 150000 155300
![]() IBA
– BAP 88670 89720 87680 164250 168000 167000
IBA
– BAP 88670 89720 87680 164250 168000 167000
Analysis of In Vivo and In Vitro proteins: The analysis of soluble proteins by SDS – PAGE methods do not show any discernable changes in the overall protein proteins profiles obtained from different tissues. On the
contrary, membrane proteins show certain distinct changes in the overall pattern, but the others though show subtle changes however are not clearly discernable. In IBA treated tissues, among many changes, the 55 - 58 KD protein were distinctly synthesized in higher quantities then in control. A similar enhanced level of same proteins is also found in BAP and IBA + BAP treated segments. However, a distinct band of l35 KD, protein found only in IBA treated tissue was district absent in other tissues. The said protein profiles were more or less identical in both membranes and in vitro translated proteins (Fig. 1 and Fig. 2).
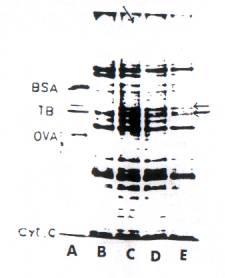
Fig. 1 SDS- polyacrylamide slab gel electrophoresis of membranes obtained from control and hormonal treated hypocotyls. Total membrane proteins were isolated from hypocotyls. The solubilized proteins were subjected to SDS-PAGE and proteins with Coomassie brilliant blue. (A) Marker protein – (B) Control; (C) IBA treated; C); IBA treated ; D) BAP treated; E) IBA – BAP treated. All hypocotyls were exposed for 12 hrs. Note the arrow in (C) shows a new protein band not found in others. Arrows against TB shows increased amount only in hormone treated segments.
DISCUSSION
Differential gene activation or inactivation in the epicotyls or hypocotyls of pea plants and the hypocotyls of soybean in response to the combined action of auxins and Cytokinins or individually, as short and long time responses, has been demonstrated by the methods of differential screening of C-DNA clones and also by the analysis of in vitro translated radioactive proteins by 2-Dimensional gel electrophoresis (Theologis, 1982; Keinschmidt, 1984; Theologis, 1986; Kuhlemeir, 1987)· It is fiat accompli that redifferentiation of differentiated tissues in response to individual or the combination of hormones into specific organs encompasses differential gene activity in the target tissues entailing cell transformation into embryonic kind of meristems. Further differentiation is imposed by cell polarity fixation. The role of cytoskeleton elements including tubulins and its associated proteins such maps, ‘tau’ and other proteins in cell transformation and cell polarity fixation for directional cell division for further differentiation has been implied and such work has been extensively reviewed (Hopler, 1974; Quatrano, 1978; Lloyd, 1987)·
The excised hypocotyl segments of French beans have been used as beau ideal system for understanding the roles and interplay of auxins and cytokinin in the induction or repression of new root formation. The IBA at the concentration of 5 x 10 -15M or more induces maximal number of roots, but BAP at the concentration of 1.5 x 10-6 M or less totally inhibits IBA induced root initiation. However the BAP at a concentration of 1.5 x 10^6 m or less, with IBA enhances auxin medicated root formation which is not significant (data not given) the observation of which adduces that adventitious root formation or its inhibition is dependent on the balance between the auxin and cytokinin concentration ratio. In all our experiments, inhibitory concentration of BAP (4.5 x 10-6 M) is used and the ratio between them is consistently maintained.
Anatomical and cytological studies luculently affirm that the pericycle cells located in between split exarch xylem act as target cells for IBA's potentation into new roots. But cytokinins at higher concentrations inhibit the transformation of IBA's target tissue into new root initials; instead the target tissue and its adjunct cells get activated into undifferentiated proliferating kind of cells. The quintessential features of cytokinin's stymied action is that it is effective only if auxin treated segments are exposed to BAP wall before 12 hrs, but at later time, it fails to abolish auxin induced root formation. This avowedly proffers, whatever molecular events actuated in the form of gene activation or repression, within the said time frame are critical and as well as potentation events for root initiation or inhibition.
To adduce differential gene activity in response to hormones, studies on changes in the pattern or protein synthesis pari passu RNA synthesis in time-frame starting from 30 mins to 24 hrs, obviously imply and infer that both IBA and BAP independently and in combination enhance the rate of protein synthesis very early (30 mins) without augmenting or activating transcriptional activity. So the hormonal activation of increased protein synthesis can be adduced to the activation of protein synthesis machinery. The auxin alone, however, potentates enhanced transcriptional activity entailing the production of higher amounts of RNA pari passu poly(A)+RNA at 2 hrs and onwards, but BAP alone, inspite of having no effect on tissue into any apparent morphological manifestations, enhances transcriptional activity only after 6 hrs of treatment. On the contrary, BAP represses IBA activated early (at 2 hrs) transcriptional activity and maintains its repression till 6 hrs, which proffers that BAP's repression is transitory but regulatory. Not withstanding, the enhanced protein synthesis at later periods i.e. 12 to 24 hrs n hormone treated segments is bolstered by increased rate of transcriptional activity. Determination of the rate of total RNA synthesis (Table 2) specific activity of poly (A) +RNA synthesis (data not given) and quantification of template activity of total RNA (Table3) by the way of in vitro translation, affirm that mRNA content increases in aIl hormone treated segments over 24 hrs of time by two to four fold over control tissues·
Analysis of both in vivo and in vivo synthesized proteins derived from 24 hrs stage of segments by SDS~-PAGE methods have been used to determine differentiate Cytokinin’s stymied action on IBA induced gene activated or its represses products. Protein profiles of soluble proteins (105 x G supernatant fraction) don't show any distinctly identifiable changes in the overall pattern. Among the mélange of protein profiles obtained from in vivo and in vitro methods, in spite of many subtle and not so clearly discernable changes found in the overall pattern, the most distinct and delineative changes among the pot pouri radioactive protein profiles obtained from in vitro translated poly (A) +RNA products (Fig· 2) which matched exactly with the in vivo membrane protein profiles (Fig· 1) are identified. And their molecular weights have been determined as 55 - 58 Kd and 135 Kd by using standard proteins as markers (on a semi-log graph).
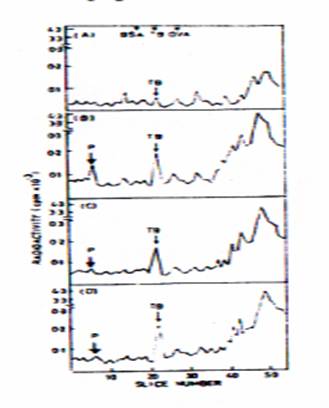
Fig· 2. Radioactive profile of in vitro translated poly(A) +RNA products obtained from control and hormone treated hypocotyls at 12 hrs· The translated product were separated by SDS-PAGE method· The gels were sliced into 1 mm pieces, digested with H202 and the radioactivity was measure · (A) Radioactive protein profiles from control segments; (B) IBA treated segments; (C) BAP treated segments; (D) IBA + BAP treated segments· BSA(Bovines Serum Album)TB (Tubulins), OVA(Ovalbumin), Cyt.C (Cytochrome C) and P(Putative protein. Note the newly induces putative "P" protein in (B) and its absence in (C) and (D), and also note increased amounts in TB only in hormone treated segments and not in control
Substantial increase in 55-58 KD proteins only in those segments treated with different hormones proffers that both IBA and BAP independently or in combination, potentate and enhances the transcriptional activity of 55-58 KD protein -encoding genes. In earlier studies (Kantharaj et al., 1985), the 55-58 KD proteins have been identified as a and b tubulins by immunological and other methods though the requirement to tubulins in higher quantities can be
accounted for IBA and IBA + BAP treated tissues for their root initiating and cell proliferating activities, their requirement in BAP tissues is shrouded with enigma for the tissue doss not manifest in any anatomical or cytological changes·
In contrast to tubulins, the novel & centre temp 135 KD protein band distinctly found only in IBA treated segments is unequivocally marked by its total absence in other tissues. This affirms that the putative protein is an induced gene product ostensibly activated by IBA. The recreation of the said protein by cytokinin suggests that cytokinin plays a regulatory role on auxin-activated gene. However, the 135 KD protein's role and character could not be ascertained at this joint of time except for the fact that both 55 - 58 KD and 135Kd proteins are associated with membranes. Tubulins and their associated high molecular weight proteins have been implicated to act as nucleating centers in the membranes and are also involved in the orientation of mitotic spindle fibers in the mitotically transformed cells (Hopler, 1974; Quatrano, 1978). Whether or not the IBA induced gene products have any such role in root initiation and whether Cytokinins play a regulatory role in inhibiting it or not, is still a profoundly esoteric phenomenon to be expedited·
In an avowed effort to delineate molecular events that regulate IBA induced root initiation and its inhibition by the regulatory activity of cytokinin, C-DNA clones have been prepared against different mRNA populations obtained from different tissues and the screening process is in progress·
ACKNOWLEGEMENT
G. R· Kantharaj sincerely acknowledges, with a deep sense of gratitude, to Prof· G. Padmanabhan, for being a lodestar and a friend in my research activities.
REFERENCES:
1. Arnold, D· and Joseph, A (1968) in Experimental Physiol, (Holt Rinehart dc Winston, London. ed.), pp· 265-268.
2. Davies, 's. and Larkins, H. (1973), Plant Physiol., 52, 339-343·
3. Hagen, G et al. (1984), Planta, 162, 147-153.
4. Hopler, P·H & Palevitz, S·A (1974), Ann· Rev. Plant Physiol., 25, 309-316.
5. Kantharaj, G.R et al. (1979), Phytochemistry, 18, 383-387·
6. Kantharaj, G.R et al. (1985), Phytochemistry, 24, 23-27·
7· Kuhlemeier, C ET el., (1987), Ann. Rev. Plant Physiol 38, 221-257·
8. Laemli, U.R (1970), Nature (London), 227, 680-d82.
9. Lloyd, C.W (1987), Ann· Rev. Plant Physiol., 38, 119-139·
10. Lowry, O.H et al., (1957), J· Biol. Chem., 19j, 265-270.
11. Miller, C.0 (1961), Ann. Rev Plant Physiol., 12, 259-267.
12. Padmanabhan, G et al., (1975), Proc. Nat. Acad. Sci·, USA., 72, 4293-4297·
13. Pennman, 3 (1996), J.Mol. Biol., 17, 117-122
14. Quatrano, R.S (1978), Ann· R3v. Plant Physiol., 29, 487-494.
15. Roberts, 9·3 dc Patterson, H.M. (1973)· Proc. Nat. Acad. Sci., Sci., U.S.A 70, 233-237·
16. Rosen, H (1957), Arch. Biochem· Biophys·, 67, 10-14.
17· Scott, M.P (1985), Tends in Genetics, 1, 74-80.
18. Theologis, A de Ray, P.M (1982), Proc. Nat. Acad· Sci., U.S.A., 79, 418-421.
19· Theologis, A (1986), Ann. Rev. Plant Physiol., 37, 407-438.
20. Tarvis, R·L et al.,:(1973), Plant Physiol., 52, 608-61u.
21.'Travis, R·L dc Key, ;J.L (1976), Plant Physiol., 57, 936-940·
22. Trewavas, A.J (1976), in Molecular aspects of Gene expression in plants, (Bryant J.A., ed.), pp. 249-259, Academic Press, NY, USA.
J. Prakash and R. L. M. Pierik (eds.), Horticulture - New Technologies and Applications. 131-139.
Ó1991 Kluwer Academic Publishers. Printed in the Netherlands.